
AG129小鼠模型在蟲媒病毒感染機制和機體免疫反應研究中的應用
節肢動物媒介病毒(蟲媒病毒)是影響熱帶和亞熱帶地區約39億人公共衛生的主要威脅[1]。在生態、經濟和社會因素共同作用下,這些病毒在全球爆發的頻率和規模不斷增加。登革病毒(DENV)和黃熱病毒(YFV)數百年來一直威脅著人類健康,近年來,寨卡病毒(ZIKV)和基孔肯雅病毒(CHIKV)等新發病毒進一步加劇了公共健康負擔。更為重要的是,大多數蟲媒病毒尚無特效療法或有效疫苗。因此,蟲媒病毒的預防和治療一直是科學研究的重點方向。
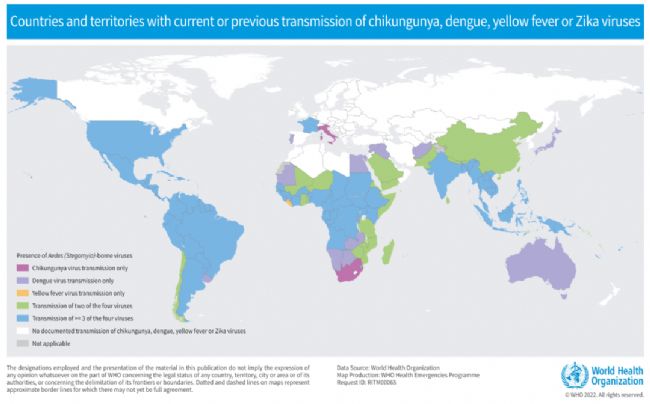
圖1 主要的蟲媒病毒全球分布情況[1]
免疫功能健全小鼠大多對蟲媒病毒不敏感
干擾素(IFN)是抵御病毒感染的第一道防線中的強效細胞因子,通過直接誘導可抑制病毒復制的抗病原體分子,實現炎癥和免疫控制。蟲媒病毒編碼特定蛋白以拮抗干擾素(IFN)反應,從而對抗宿主免疫。高效的IFN拮抗作用對蟲媒病毒至關重要,因為這些病毒需要在血液中保持足夠的病毒載量(病毒血癥)以維持其病媒-宿主循環[2]。然而,這類IFN拮抗作用在小鼠中似乎不存在,因此免疫功能健全的小鼠對大多數蟲媒病毒并不敏感[3-5]。例如,登革病毒(DENV)的非結構蛋白NS5通過靶向降解人類IFN下游轉錄激活因子STAT2來抑制Ⅰ型IFN反應,從而在人體中實現復制并引起疾病,但小鼠STAT2蛋白對NS5具有抵抗性,因而無法成功感染[5-6]。因此,多種IFN信號途徑分子缺陷的免疫功能低下小鼠被用于蟲媒病毒研究。
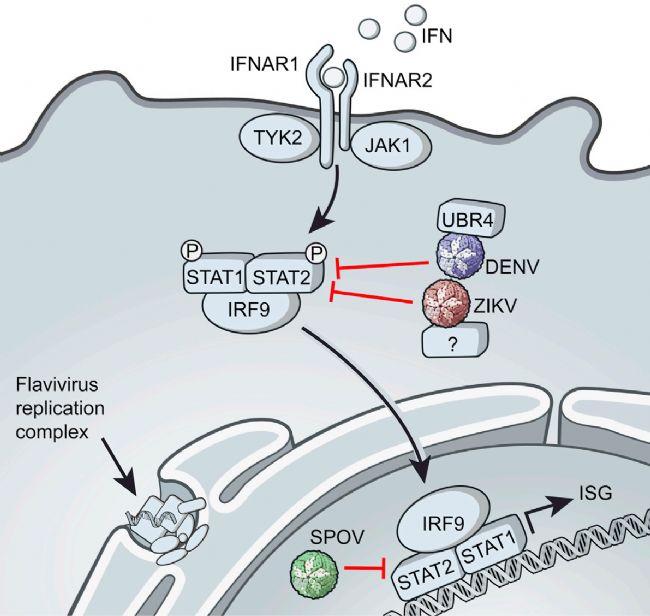
圖2 登革熱病毒(DENV)和寨卡病毒(ZIKV)通過靶向人類STAT2來抑制Ⅰ型IFN信號傳導[5]
129背景的AG129小鼠被廣泛用于蟲媒病毒和IFN通路研究
研究表明,與C57BL/6和CD-1等品系相比,129品系小鼠對部分蟲媒病毒更敏感,盡管接種后不出現癥狀,但可以在血液、卵巢和脾臟中檢測到病毒,更適合用于評估此類病毒的致畸效應[7-8]。此外,129品系也是研究IFN信號傳導通路缺陷的常用品系,特別是在與病毒感染相關研究中,目前大多數用于蟲媒病毒研究的IFN通路相關模型都是129品系[9-10]。129品系的IFNα/β/γR KO小鼠(即AG129小鼠)作為Ⅰ型(α/β)干擾素受體(Ifnar1)和Ⅱ型(γ)干擾素受體(Ifngr1)雙基因敲除模型,對蟲媒病毒極為敏感,感染后容易產生與人相似的病毒血癥,被廣泛用于這些病毒的感染研究[2-3]。最近研究表明,AG129小鼠可作為研究包括DENV、ZIKV、YFV和CHIKV在內的幾乎所有蟲媒病毒感染機制、免疫反應、預防疫苗和潛在治療方法的綜合模型[11]。

圖3 AG129小鼠是研究蚊子向脊椎動物傳播各類蟲媒病毒的有效模型[12]
賽業生物通過將129品系背景的Ifnar1 KO (A129)小鼠(產品編號:I001199)和Ifngr1 KO (G129)小鼠(產品編號:I001200)配種,構建了AG129小鼠模型(產品編號:I001125)。兩種單基因敲除小鼠體內Ifnar1基因或Ifngr1基因表達被完全敲除,且存在輕度的免疫缺陷表型。純合AG129小鼠發育正常且可育,可用于蟲媒病毒感染機制和機體免疫反應研究,以及預防性疫苗和潛在療法研發。以下是129品系的Ifnar1 KO (A129)小鼠和Ifngr1 KO (G129)小鼠基因表達和免疫細胞檢測數據。
Ifnar1和Ifngr1基因表達檢測
Ifnar1 KO (A129)小鼠大腦和十二指腸中Ifnar1基因表達被完全敲除,Ifngr1 KO (G129)小鼠十二指腸、胸腺和腦組織中Ifngr1基因表達同樣被完全敲除。
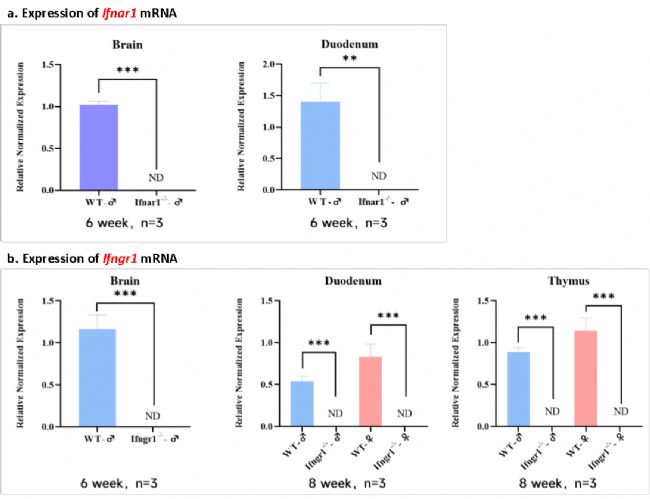
圖4 Ifnar1 KO (A129)小鼠和Ifngr1 KO (G129)小鼠體內Ifnar1和Ifngr1基因表達的RT-qPCR檢測
淋系細胞亞型比例檢測
與野生型小鼠相比,Ifnar1 KO (A129)小鼠和Ifngr1 KO (G129)小鼠均表現為外周血T細胞比例升高,而B細胞和NK細胞比例降低的表型。

圖5 Ifnar1 KO (A129)小鼠和Ifngr1 KO (G129)小鼠外周血中T/B/NK細胞流式細胞術檢測
髓系細胞亞型比例檢測
與野生型小鼠相比,Ifnar1 KO (A129)小鼠和Ifngr1 KO (G129)小鼠的外周血全髓系細胞(CD11b+)比例均略有降低,粒細胞、巨噬細胞和樹突狀細胞比例無顯著性差異。
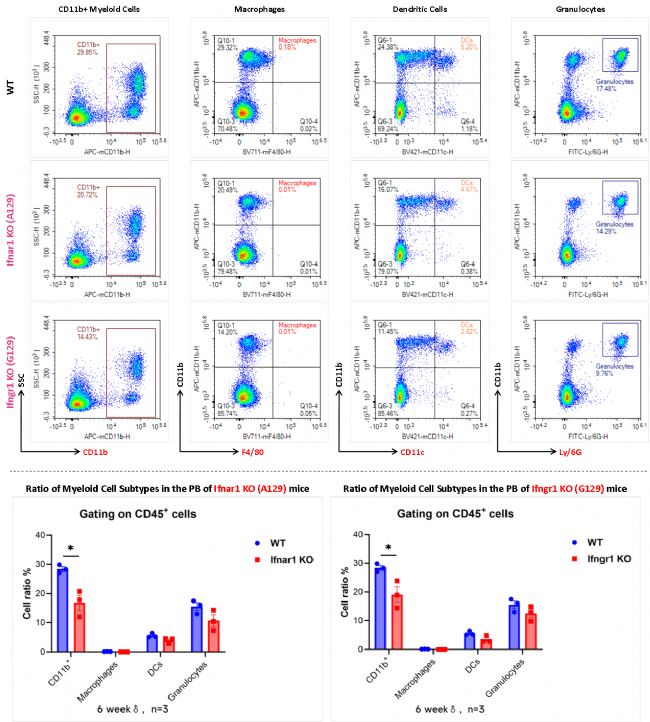
圖5 Ifnar1 KO (A129)小鼠和Ifngr1 KO (G129)小鼠外周血中各髓系細胞亞型流式細胞術檢測
總 結
賽業生物通過將129品系背景的Ifnar1 KO (A129)小鼠(產品編號:I001199)和Ifngr1 KO (G129)小鼠(產品編號:I001200)配種,構建了AG129小鼠模型(產品編號:I001125)。純合AG129小鼠可用于蟲媒病毒感染機制和機體免疫反應研究,以及預防性疫苗和潛在療法研發。
此外,賽業生物還提供多種免疫缺陷品系和人源化模型,以滿足研發人員在病毒感染和免疫相關研究領域的實驗需求。
免疫缺陷疾病模型推薦
參考文獻:
[1]World Health Organization. (2022, March 31). Global Arbovirus Initiative. Retrieved July 10, 2024, from
https://www.who.int/initiatives/global-arbovirus-initiative
[2]Müller U, Steinhoff U, Reis LF, Hemmi S, Pavlovic J, Zinkernagel RM, Aguet M. Functional role of type I and type II interferons in antiviral defense. Science. 1994 Jun 24;264(5167):1918-21.
[3]van den Broek MF, Müller U, Huang S, Zinkernagel RM, Aguet M. Immune defence in mice lacking type I and/or type II interferon receptors. Immunol Rev. 1995 Dec;148:5-18.
[4]Marín-Lopez A, Calvo-Pinilla E, Moreno S, Utrilla-Trigo S, Nogales A, Brun A, Fikrig E, Ortego J. Modeling Arboviral Infection in Mice Lacking the Interferon Alpha/Beta Receptor. Viruses. 2019 Jan 8;11(1):35.
[5]Grant A, Ponia SS, Tripathi S, Balasubramaniam V, Miorin L, Sourisseau M, Schwarz MC, Sánchez-Seco MP, Evans MJ, Best SM, García-Sastre A. Zika Virus Targets Human STAT2 to Inhibit Type I Interferon Signaling. Cell Host Microbe. 2016 Jun 8;19(6):882-90.
[6]Ashour J, Morrison J, Laurent-Rolle M, Belicha-Villanueva A, Plumlee CR, Bernal-Rubio D, Williams KL, Harris E, Fernandez-Sesma A, Schindler C, García-Sastre A. Mouse STAT2 restricts early dengue virus replication. Cell Host Microbe. 2010 Nov 18;8(5):410-21
[7]Lazear HM, Govero J, Smith AM, Platt DJ, Fernandez E, Miner JJ, Diamond MS. A Mouse Model of Zika Virus Pathogenesis. Cell Host Microbe. 2016 May 11;19(5):720-30.
[8]Rossi SL, Tesh RB, Azar SR, Muruato AE, Hanley KA, Auguste AJ, Langsjoen RM, Paessler S, Vasilakis N, Weaver SC. Characterization of a Novel Murine Model to Study Zika Virus. Am J Trop Med Hyg. 2016 Jun 1;94(6):1362-1369.
[9]Meyts I, Casanova JL. Viral infections in humans and mice with genetic deficiencies of the type I IFN response pathway. Eur J Immunol. 2021 May;51(5):1039-1061.
[10]Zivcec M, Spiropoulou CF, Spengler JR. The use of mice lacking type I or both type I and type II interferon responses in research on hemorrhagic fever viruses. Part 2: Vaccine efficacy studies. Antiviral Res. 2020 Feb;174:104702.
[11]World Health Organization. "Status of Preclinical Model Development for Zika Disease, Including Challenge and Pregnancy Models." WHO, Accessed 2 Aug. 2024, From:
[12]Baldon LVR, de Mendonça SF, Ferreira FV, Rezende FO, Amadou SCG, Leite THJF, Rocha MN, Marques JT, Moreira LA, Ferreira AGA. AG129 Mice as a Comprehensive Model for the Experimental Assessment of Mosquito Vector Competence for Arboviruses. Pathogens. 2022 Aug 3;11(8):879.



