
病毒感染標志dsRNA(雙鏈RNA)的形成機制及生物學意義
dsRNA的形成機制
第一種機制:基于T7 RNA聚合酶(T7 RNAP)本身具有的RNA依賴性RNA聚合酶(RdRp)活性,能夠以產物RNA為模板,3'末端繼續延伸RNA后與原RNA分子反向互補配對形成發卡結構;或者是在轉錄過程中,隨機斷裂的/不完整的轉錄產物與mRNA互補配對從而延伸成不完整的雙鏈RNA。
第二種機制:以DNA反義鏈為模板,獨立于啟動子的轉錄過程形成dsRNA
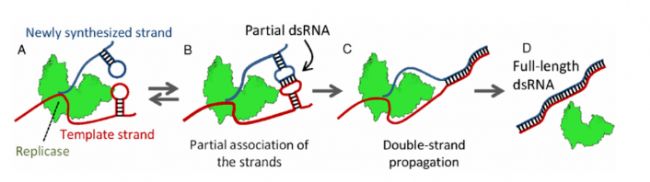
圖1 dsRNA復制示意圖
dsRNA經典克隆
J2克隆
J2單克隆抗體(Anti-dsRNA mAb J2)是dsRNA檢測的金標準,具有高度特異性。它最初用于植物病毒的研究,但自從Weber等人在2006年的開創性論文發表以來,J2被用來證明所有正鏈RNA病毒在感染的細胞中產生的大量dsRNA。
K2克隆
K2單克隆抗體(Anti-dsRNA mAb K2)是另一種抗dsRNA的單克隆抗體,主要用于ELISA和夾心法ELISA實驗,K2主要用于ELISA和夾心法ELISA,在高背景環境中仍能精準識別dsRNA,尤其在高濃度L-dsRNA和長時間孵育條件下,顯示出對天然dsRNA結合能力最強的特性。
dsRNA的生物學意義
1. dsRNA通常被視為病毒感染的標志,因為許多病毒(尤其是雙鏈RNA病毒)以dsRNA作為其基因組或復制中間體。例如正鏈RNA病毒(+ssRNA)在復制時會先合成負鏈RNA(-ssRNA),形成dsRNA,然后再從負鏈RNA合成更多的正鏈RNA。
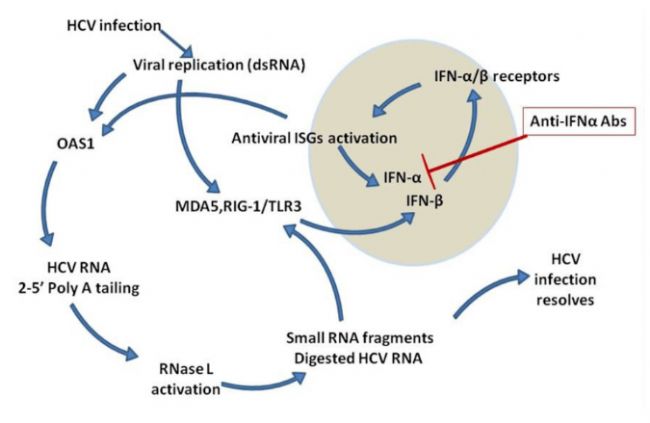
圖2 病毒感染后產生的dsRNA被RIG-I、MDA5和TLR3等受體識別,觸發IFN-β的產生,進而激活ISGs,包括OAS和IFN-α,形成抗病毒反應的正反饋循環。抗IFN-α抗體中和IFN-α,抑制ISGs的誘導,削弱免疫反應,影響病毒清除。
2. 在mRNA疫苗和治療中,dsRNA作為副產物在體外轉錄(IVT, In Vitro Transcription)制備mRNA過程中產生,能夠被細胞內的核酸受體識別,例如TLR3、MDA5或者RIG-I,進而引發天然免疫炎癥反應。
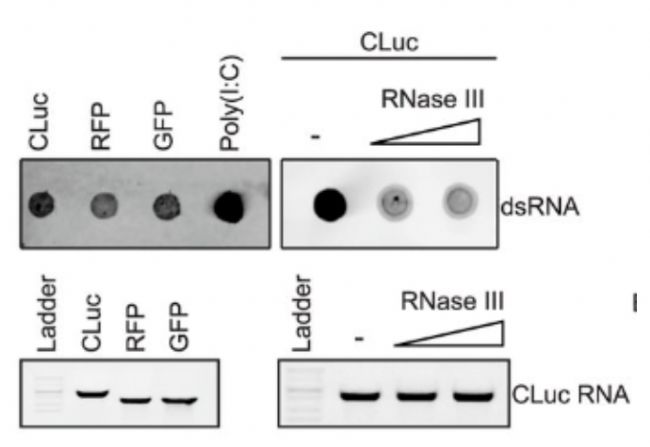
圖3 在 IVT 反應過程中形成 3′-延伸的 RNA 副產物。(A) 使用 dsRNA 特異性抗體 (J2) 對三種不同的 mRNA 序列(CLuc、RFP 和 GFP)進行 dsRNA檢測。
3. 免疫信號通路中,dsRNA作為重要的病原分子模式(PAMPs),能夠激活天然免疫系統中的關鍵受體(如RIG-I、MDA5和TLR3)。這些受體通過識別dsRNA,引發干擾素(IFN)等抗病毒分子的產生,從而觸發強大的免疫反應。
dsRNA結合后,NLRP1觸發其UPA和CARD結構域的釋放,組裝炎癥體種子,誘導炎癥體形成并激活caspase 1。激活的caspase 1切割炎癥細胞因子前體(如IL-1β和IL-18)和一個形成孔洞的蛋白gasdermin D(GSDMD)。GSDMD孔洞在質膜上形成,誘導焦亡,這是一種炎癥形式的細胞死亡。
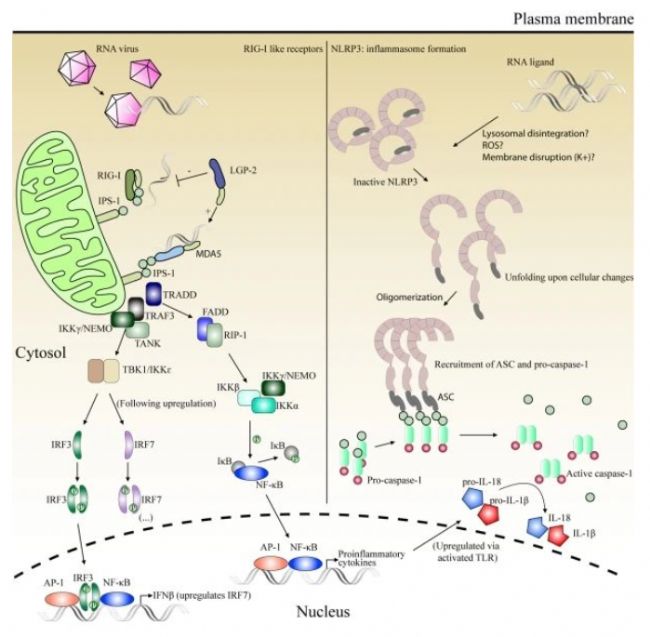
圖4 dsRNA作為觸發RIG-I(Retinoic Acid-Inducible Gene I)和MDA5(Melanoma Differentiation-Associated Protein 5)識別的分子模式,從而啟動先天免疫反應。
dsRNA不僅在理解病毒感染機制和宿主免疫反應中扮演著關鍵角色,而且在開發新的抗病毒策略和疫苗技術方面具有重要價值。通過對dsRNA的形成機制、識別受體以及其在免疫激活中的作用的深入研究,科學家們能夠更好地理解病毒如何逃避或操縱宿主的天然免疫反應,以及如何設計更有效的疫苗和治療藥物來對抗病毒感染。
兔單克隆抗體優勢
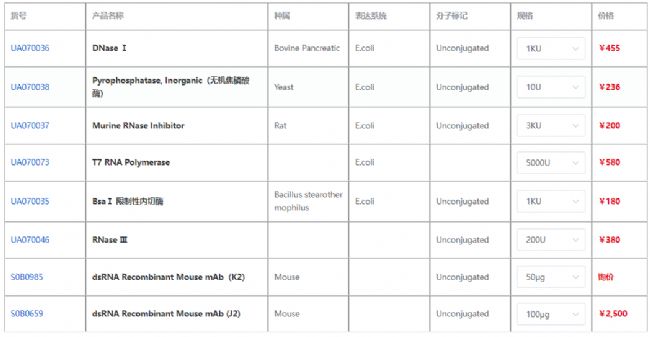
斯達特上架抗dsRNA抗體,引用經典克隆J2和K2,高批間一致性和高靈敏度;還有更多相關的高純度、高活性蛋白,為廣大客戶提供高品質抗體蛋白。
參考文獻
[1] Jensen S, Thomsen AR. Sensing of RNA viruses: a review of innate immune receptors involved in recognizing RNA virus invasion. J Virol. 2012 Mar;86(6):2900-10. doi: 10.1128/JVI.05738-11. Epub 2012 Jan 18. PMID: 22258243; PMCID: PMC3302314.
[2] Bharti, Praveen & Kumar, Vijay & Gumma, Phani & Karna, Rahul & Kar, Premashis. (2020). Can We Predict the Response to Interferon: A Therapy in Chronic Hepatitis C Patients Using Anti-Interferon-A Antibody Levels?. Tropical gastroenterology: official journal of the Digestive Diseases Foundation. 22-29.
杭州斯達特(www.starter-bio.com)志在為全球生命科學行業提供優質的抗體、蛋白、試劑盒等產品及研發服務。依托多個開發平臺:重組兔單抗、重組鼠單抗、快速鼠單抗,重組蛋白開發平臺(E.coli,CHO,HEK293,Insect Cells),已正式通過歐盟98/79/EC認證、ISO9001認證ISO13485。



