
JAK-STAT 通路的組成激活與炎癥和自身免疫性疾病應用
JAK-STAT 信號通路在進化上是保守的。它由配體 (細胞因子)-受體復合物、JAKs 和 STATs 組成。JAK 家族的 4 個成員分別為 JAK1-3 和 TYK2。STAT 家族成員分別為:STAT1, STAT2, STAT3, STAT4, STAT5 (STAT5A 和 STAT5B) , 和 STAT6。
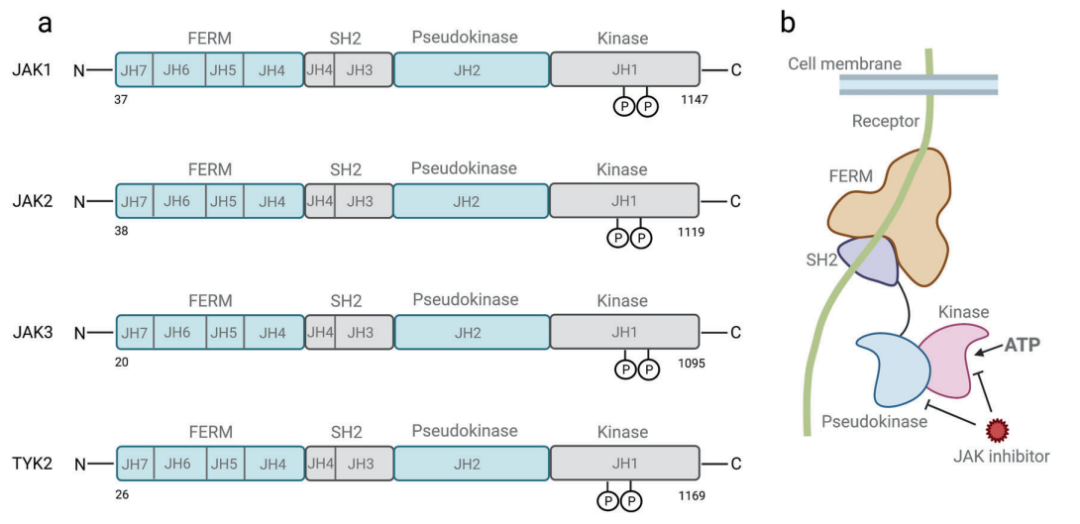
圖 1. JAK 的結構和 STAT 蛋白家族的結構[1]
經典的 JAK-STAT 信號傳導始于細胞因子與其相應的跨膜受體之間的細胞外結合。兩個細胞因子受體家族,I 型和 II 型 (type I and type II receptors),是JAK-STAT 信號通路上的兩類重要受體。
細胞/生長因子與相應的受體結合,導致對應受體二聚化和相關 JAK 的募集,進而促進 JAK 激活;活化的 JAK 導致結合受體酪氨酸磷酸化、形成 STATs 的對接位點。
接著,JAK 介導 STATs 酪氨酸磷酸化 (p-Tyr),然后 STATs 從受體上解離形成同源二聚體或異源二聚體。同源二聚或異源二聚的 STATs 易位至靶基因啟動子,調節靶基因的轉錄。
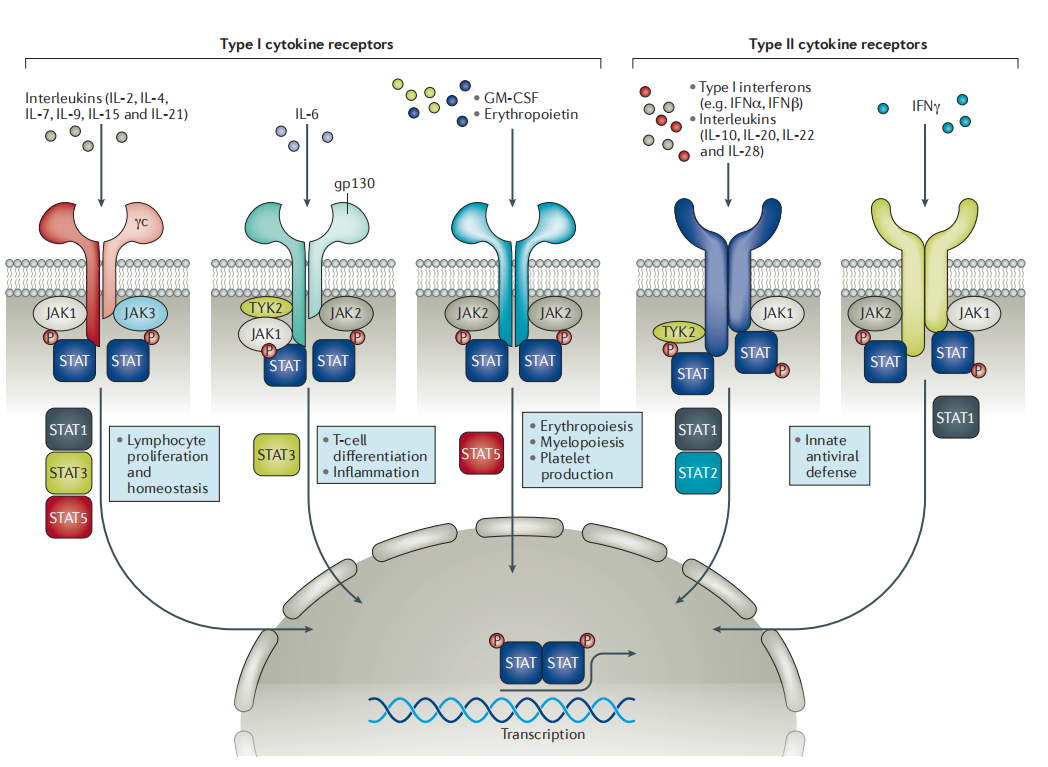
圖 2. JAK 家族介導的細胞內信號通路[3]
為了維持 JAK-STAT 通路的平衡和穩定狀態,許多負調節因子參與了 JAK-STAT 信號轉導的調節。這些負調節因子分為三種類型:活化 STAT 蛋白抑制因子 (PIAS)、SOCS/CIS 家族成員和蛋白酪氨酸磷酸酶 (PTPs)。
PIAS:PIAS 主要與 STAT 二聚體相互作用,抑制 STAT 與 DNA 結合,從而阻斷 JAK-STAT 信號轉導。
CIS/SOCS 家族:通過三種方式負向調節 JAK-STAT 通路:(1) 與酪氨酸激酶受體結合,阻斷 STAT 的募集;(2) 直接與 JAK 結合,抑制其激酶活性;(3) SOCS 蛋白結合 JAK 或 STAT,結合的 SOCS 蛋白與延長蛋白 B/C-cullin5 復合物相互作用,然后通過多泛素化和蛋白酶體降解 JAK 或 STAT。
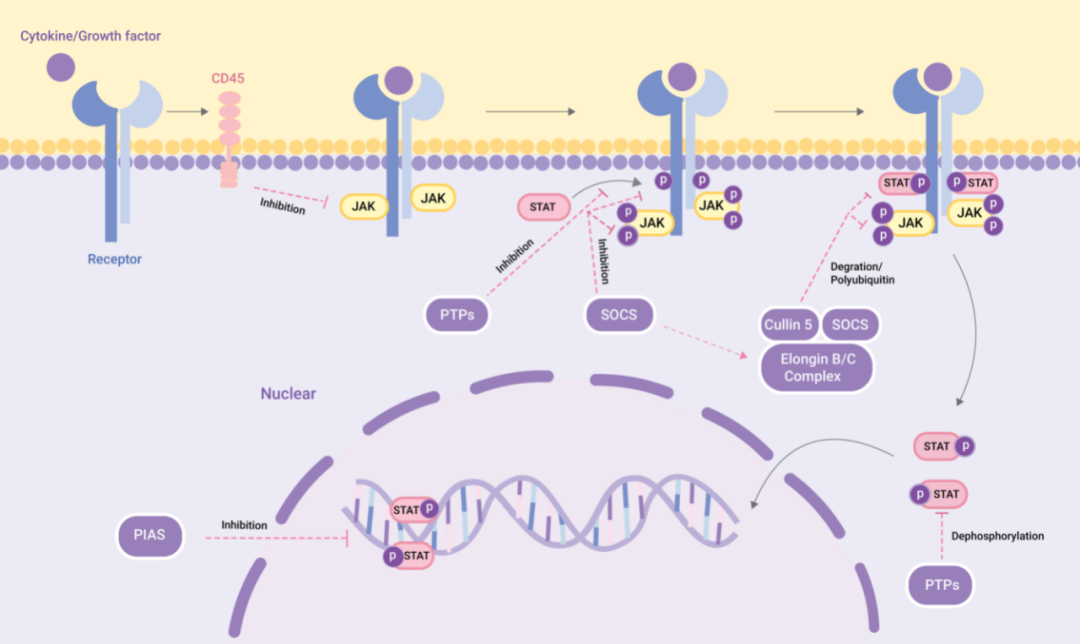
圖 3. 經典 JAK-STAT 信號通路的激活和負調節[1]
PTPs:通過與 JAK、STAT 或受體相互作用來抑制 JAK-STAT 通路: (1) 使 STAT 二聚體去磷酸化;(2) 與受體相互作用,使相關的 JAK 去磷酸化;(3) 在 CD45 (一種跨膜 PTP) 的情況下,抑制 JAK 的磷酸化。
在某些病理狀況下,炎癥過程被認為是身體受到病原體攻擊或損傷時的局部保護反應。JAK-STAT 是由細胞因子調節的主要信號通路,對于啟動先天免疫、協調適應性免疫機制以及最終抑制炎癥和免疫反應至關重要。
類風濕性關節炎 (RA) 是一種全身性、多關節的慢性、炎癥性的滑膜關節肌肉骨骼疾病。與 RA 相關的大量組織損傷可發生在心臟,以及肺、腎臟和血管等。許多參與 RA 的細胞因子 (尤其是 IL-6、IFN-γ、IL-12/23) 通過 JAK-STAT 途徑發出信號。
炎癥性腸病 (IBD) 是一種慢性或復發性胃腸道炎癥性疾病,表現為腸道內穩態失調,其特征是黏膜免疫系統不受控制的炎癥和異常激活。研究認為 IBD 是由多種因素引起的,包括遺傳易感性、環境因素以及腸道菌群和免疫系統功能的變化。同樣,許多細胞因子通過其促炎或抗炎作用在 IBD 的病理生理過程中發揮關鍵作用,因此,對 JAK/STAT 信號通路是 IBD 的一個有吸引力的潛在治療靶點。
JAK 抑制劑 (JAKinibs) 在治療包括 RA 在內的幾種炎癥性疾病中顯示出良好的前景。第一代 Jakinibs 如 Tofacitinib,Baricitinib 和 Ruxolitinib 已經被 FDA 批準用于自身免疫疾病。然而,第一代 Jakinibs 的非選擇性 (pan-JAK) 作用可能與不良副作用有關,下一代 Jakinibs 如 Decernotinib,GLPG0634,Upadacitinib 等則提高了選擇性。然而,增加選擇性也可能降低功效。此外,幾家生物技術公司正在開發 JAK 抑制劑,目標是創造一種具有最大功效和最小脫靶效應的分子,如 SHR0302,Abrocitinib,Delgocitinib 等。
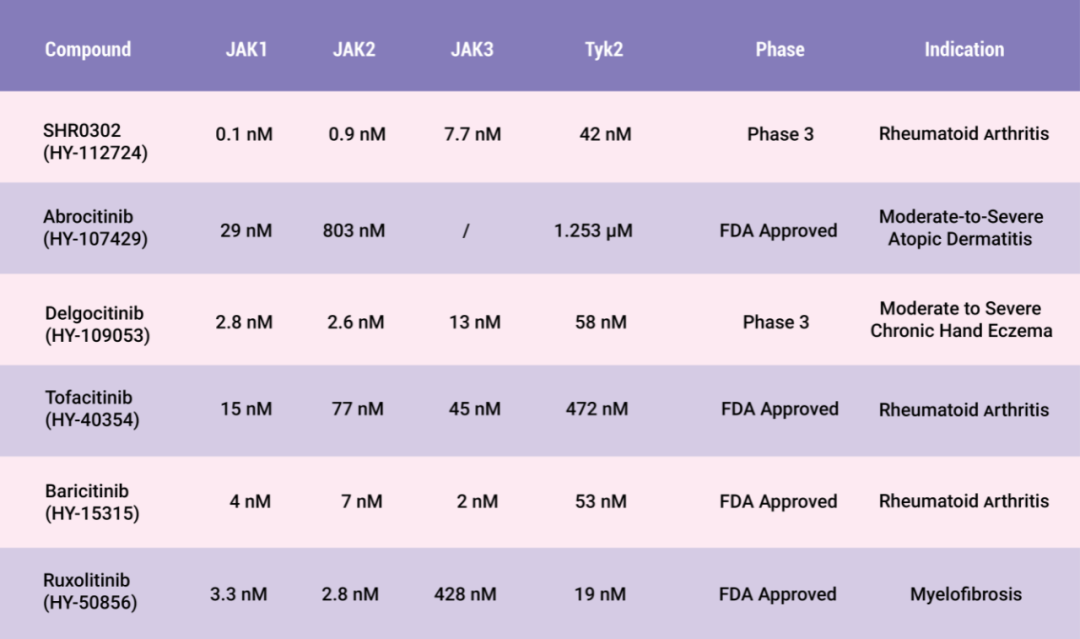
圖 4. 部分 Jakinibs 特點
作為 I/II 型細胞因子受體下游的 JAK 底物和關鍵信號分子,STATs 已被研究為治療炎癥和自身免疫以及惡性腫瘤的有吸引力的靶標。如 STAT3 對于 IL-6 下游的信號傳導至關重要 (IL-6 調節 T 細胞和其他免疫細胞產生 IL-17,在風濕病的發病機制中起重要作用)。此外,已在一些人類癌癥和癌細胞系中觀察到 STAT3 和 STAT5 的組成型激活。
但是,STAT 抑制劑開發面臨的挑戰包括生物利用度和選擇性問題。例如,STAT1 和 STAT3 之間存在相當大的同源性;不同 STAT 的功能冗余等。但仍有 STAT 抑制劑進入臨床,如 OPB-31121 和 OPB-51602 (STAT3 抑制劑)。
我們相信隨著新數據的出現和新藥的繼續開發,可能很快還會有更多的 Jakinibs 出現。在未來幾年,Jakinibs 也有望改變自身免疫和風濕病領域,因為正在進行的臨床前和臨床相關研究肯定會確定選擇性、優化劑量、配方等可行性和益處,以最大限度地減少不良脫靶,并提高臨床療效。
|
相關產品 |
|
具有口服活性的 JAK3/2/1 抑制劑,IC50 分別為 1, 20 和 112 nM。 |
|
選擇性的,具有口服的 JAK1 和 JAK2 抑制劑,IC50 分別為 5.9 nM 和 5.7 nM。 |
|
選擇性的 JAK1/2 抑制劑,IC50 值分別為 3.3 nM 和 2.8 nM,選擇性是 JAK3 的 130 多倍;誘導自噬,通過毒性線粒體自噬殺死腫瘤細胞。 |
|
有效的,具有口服活性的 JAK3 抑制劑,對 JAK3, JAK1, JAK2 和 TYK2 的 Ki 值分別為 2.5, 11, 13 和 11 nM。 |
|
選擇性的,具有口服活性的 JAK1 抑制劑,能夠有效抑制 JAK1, JAK2, JAK3 和 TYK2 的活性,IC50 值分別為 10 nM, 28 nM, 810 nM 和 116 nM。 |
|
具有口服活性的選擇性 JAK1 抑制劑 (IC50=43 nM),用于多種自身免疫性疾病的研究。 |
|
有效的具有口服活性的 JAK 抑制劑,尤其是 JAK1 的抑制劑。抑制 JAK1-STAT3 磷酸化并誘導肝星狀細胞凋亡;具有抗增殖和抗炎作用。 |
|
具有口服活性的選擇性 JAK1 抑制劑,對 JAK1 和 JAK2 的 IC50 值分別為 29 和 803 nM。抑制刺激后的 STAT1, STAT3 和 STAT5 的磷酸化水平,可用于自身免疫癥的研究。 |
|
特異性的 JAK 抑制劑,對 JAK1, JAK2, JAK3 和 Tyk2 的 IC50 值分別為 2.8, 2.6, 13 和 58 nM。 |
MCE 的所有產品僅用作科學研究或藥證申報,我們不為任何個人用途提供產品和服務。
參考文獻
1. Hu X, et al. The JAK/STAT signaling pathway: from bench to clinic. Signal Transduct Target Ther. 2021;6(1):402. Published 2021 Nov 26.2. Salas A, et al. JAK-STAT pathway targeting for the treatment of inflammatory bowel disease. Nat Rev Gastroenterol Hepatol. 2020;17(6):323-337.
3. Kevin L Winthrop, et al. The emerging safety profile of JAK inhibitors in rheumatic disease. Nat Rev Rheumatol. 2017 May;13(5):320.
4. Villarino AV, et al. Mechanisms of Jak/STAT signaling in immunity and disease. J Immunol. 2015;194(1):21-27.
5. Li WX. Canonical and non-canonical JAK-STAT signaling. Trends Cell Biol. 2008;18(11):545-551.
6. Xin P, Sun S, et al. The role of JAK/STAT signaling pathway and its inhibitors in diseases. Int Immunopharmacol. 2020;80:106210.
7. Singh S, et al. JAK-STAT inhibitors: Immersing therapeutic approach for management of rheumatoid arthritis. Int Immunopharmacol. 2020;86:106731.
8. Banerjee S, et al. JAK-STAT Signaling as a Target for Inflammatory and Autoimmune Diseases: Current and Future Prospects [published correction appears in Drugs. 2017 May;77(8):939] [published correction appears in Drugs. 2017 Jun 12;:]. Drugs. 2017;77(5):521-546.
9. Ciobanu DA, et al. JAK/STAT pathway in pathology of rheumatoid arthritis (Review). Exp Ther Med. 2020;20(4):3498-3503.
10. Kim YK, et al. Structural basis of intersubunit recognition in elongin BC-cullin 5-SOCS box ubiquitin-protein ligase complexes. Acta Crystallogr D Biol Crystallogr. 2013;69(Pt 8):1587-1597.


