
幾種 Wnt 信號通路介紹
Wnt 信號通路是一種古老的、進化上保守的通路。其中,Wnt 蛋白屬于分泌的糖蛋白,可與卷曲蛋白 (Frizzled) 家族的受體,如與脂蛋白受體相關的蛋白 5/6 (LRP5/6),Ror2 和 Ryk 相互作用,導致各種細胞內信號傳導級聯激活,這些信號傳導級聯可以交叉連接或獨立發揮作用,調節多種多樣的過程,包括細胞增殖,分化,遷移,極性和不對稱細胞分裂以及與干細胞更新有關。因此,Wnt 途徑的突變往往與人類出生缺陷、癌癥以及其他疾病有關。
細胞外 Wnt 信號刺激會引起幾種細胞內信號轉導級聯,包括 Wnt/β-catenin 依賴 (經典) 途徑和 β-catenin 獨立 (非經典) 途徑,后者又可進一步分為平面細胞極性 (PCP) 和 Wnt/Ca2+途徑。
β-catenin 依賴 (經典) 信號途徑
在有 Wnt 信號時 (如 Wnt1, Wnt3a, Wnt8),Wnt 與由 Frizzled 受體和 LRP5/6 組成的受體復合物的結合引發一系列事件,從而靶向破壞 β-catenin 所需的 APC/Axin/GSK3 復合物。具體為:Frizzled 募集的 Dishevelled (Dsh/Dvl) 導致 LRP5/6 磷酸化 (磷酸化由激酶 GSK3 和 CK1γ 介導)。磷酸化的 LRP5/6 將 Axin 募集到膜上,破壞復合物分解,從而導致細胞質中 β-catenin 的穩定和積累。隨后,β-catenin 蛋白易位至細胞核,通過取代 TLE/Groucho 復合物并募集組蛋白修飾共激活物 (如 CBP/p300,BRG1,BCL9 和 Pygo) 與 LEF 和 TCF 蛋白形成活性復合物,調節靶基因的表達。
典型的 Wnt 信號的激活可以被一些 Wnt 蛋白抑制劑抑制,如 sFRP,Dkk,WIF,Wise/SOST,Cerberus,Tiki1 等,它們的共同特征是通過阻止配體-受體相互作用或 Wnt 受體成熟來拮抗 Wnt 信號傳導。而 Wnt 信號通路也會被 R-spondin 家族蛋白和 Norrin 蛋白激活。另外,其下游途徑的激活也可以被抑制,例如 TAK1-NLK 阻斷了 β-catenin 誘導的轉錄活性。Hippo 途徑也可通過 TAZ 抑制 Wnt 信號傳導,具體為 TAZ 通過抑制 CK1δ/β 介導的 DVl 磷酸化來限制 Wnt 信號傳導。
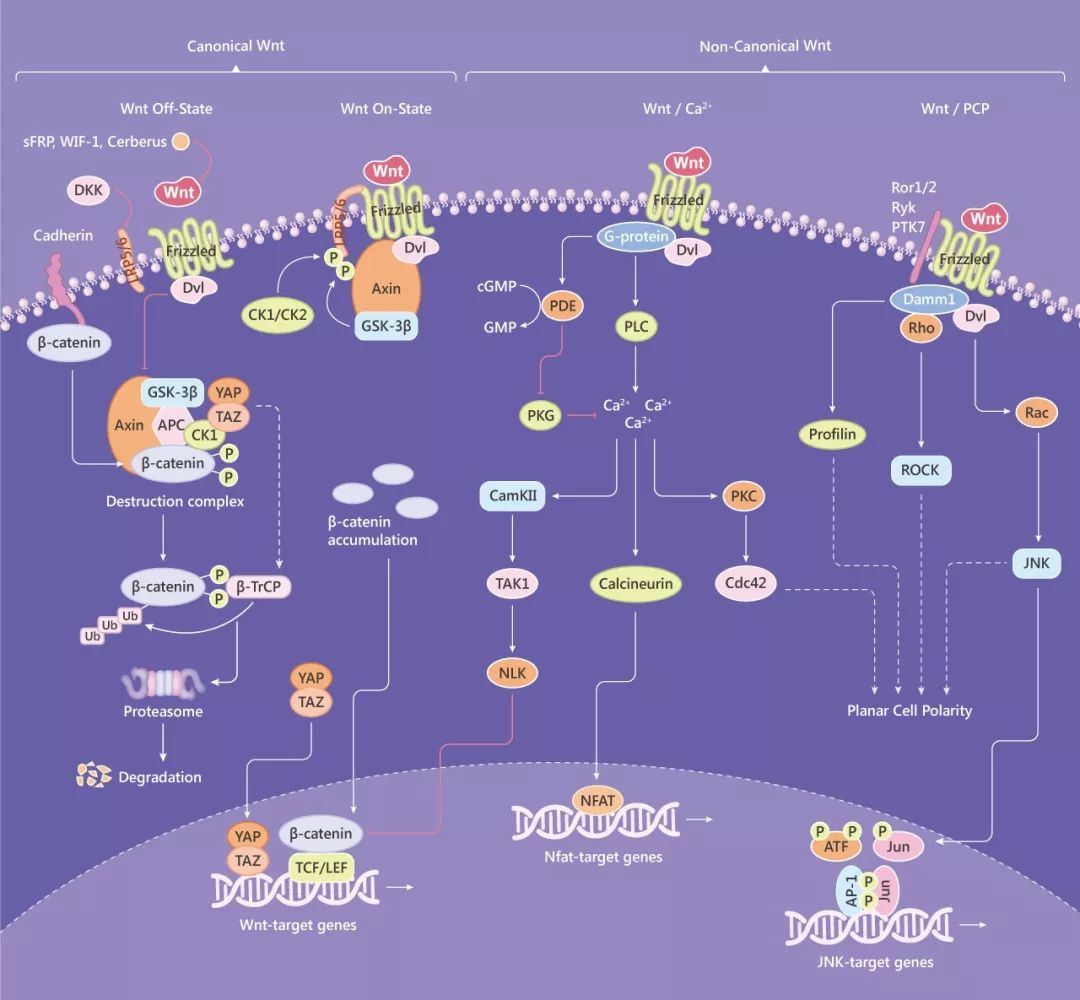 Fig. 1 典型和非典型 Wnt 信號通路圖
Fig. 1 典型和非典型 Wnt 信號通路圖
非經典平面細胞極性 (PCP) 途徑
非經典 Wnt/Ca2+途徑
Wnt抑制劑/激動劑
SalinomycinSalinomycin,一種離子載體抗生素,同時也是 Wnt/β-catenin 信號的抑制劑。Salinomycin 作用于 Wnt/Fzd/LRP 復合物,阻斷 Wnt 誘導的 LRP6 磷酸化,導致 Wnt 共受體 LRP6 蛋白降解。Salinomycin 具有較強的抗癌活性,可選擇性抑制人腫瘤干細胞。
IWP-2
IWP-2 是 Wnt 加工和分泌的抑制劑,靶向膜結合 O- 酰基轉移酶 porcupine (Porcn),Porcn 是膜結合的 O-酰基轉移酶 (MBOAT) 家族的成員,可在 Wnt 蛋白上添加棕櫚酰基,這對于其信號傳遞能力至關重要,并且是 Wnt 分泌所必需的。IWP-2 阻止 Wnt 配體棕櫚糖基化阻斷以及 Wnt 蛋白的分泌,從而抑制下游 Wnt 信號傳導。
XAV939
XAV939 是一種細胞可穿透性的 Wnt/β-catenin 信號轉導抑制劑,通過抑制聚 ADP-核糖基化酶 tankyrase1 和 tankyrase 2 來穩定破壞復合物成分 Axin,從而刺激 β-catenin 降解。
SKL2001
SKL2001 是一種通過增加 β-catenin 的穩定性來激活 Wnt 信號傳導的特異的 Wnt/β-catenin 激動劑。SKL2001 與 Axin 競爭性地與 β-catenin 的結合,破壞 Axin/β-catenin 的相互作用,從而破壞 CK1 和 GSK-3β 介導的 Ser33/37/ Thr41 和 Ser45 處 β-catenin 的磷酸化,進而增加 β-catenin 的穩定性,確保其不被蛋白酶體降解。CK1 和 GSK-3β 酶活性不受 SKL2001 影響 (如下圖)。
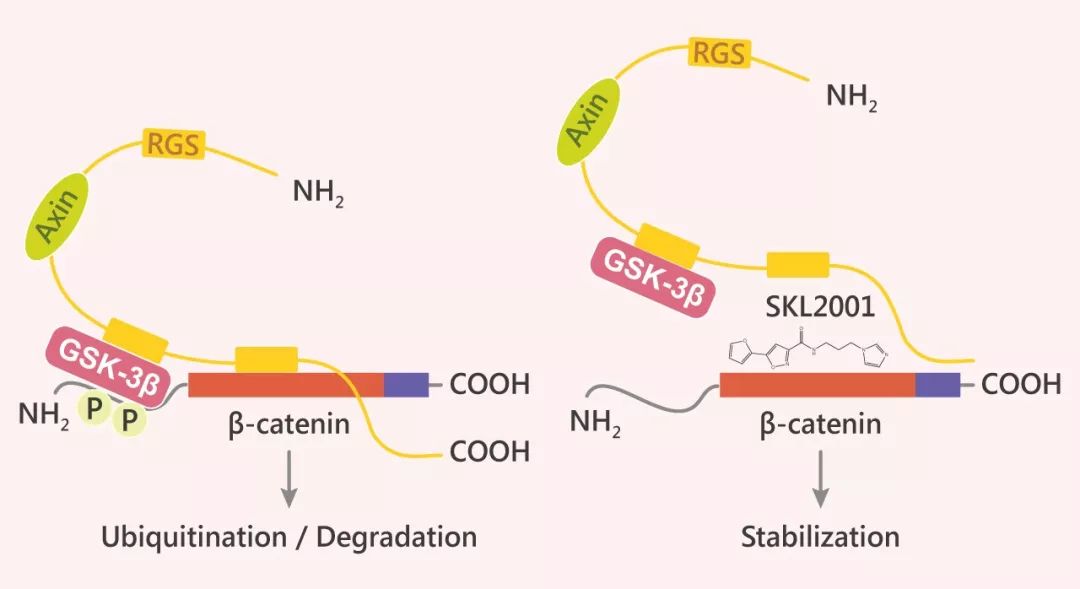
Fig. 2 SKL2001 作用機制圖
參考文獻
1. Clevers H, et al. Wnt/β-catenin signaling and disease. Cell. 2012;Jun 8;149(6):1192-205.
2. Angers S, et al. Proximal events in Wnt signaltransduction. Nat Rev Mol Cell Biol. 2009 Jul;10(7):468-77.
3. MacDonald BT, et al. Wnt/beta-cateninsignaling: components, mechanisms, and diseases. Dev Cell. 2009Jul;17(1):9-26.
4. T Zhan, etal. Wnt signaling in cancer. Oncogene. 2017 Mar;36(11):1461-1473.
5. Theka I, et al. Wnt/β-catenin signaling pathway safeguards epigenetic stability and homeostasis of mouseembryonic stem cells.
6. Komiya Y, et al . Wnt signal transduction pathways. Organogenesis. 2008 Apr;4(2):68-75.
7. Tarapore RS, et al. Modulation of Wnt/β-catenin signalingpathway by bioactive food components. Carcinogenesis. 2012 Mar;33(3):483-91.
8. Cruciat CM, et al. Secreted and transmembranewnt inhibitors and activators. Cold Spring Harb Perspect Biol. 2013 Mar 1;5(3):a015081.
9. Lien WH, et al. Wnt some lose some: transcriptional governance of stem cells by Wnt/β-catenin signaling.Genes Dev. 2014 Jul 15;28(14):1517-32.
10. Tang QL, et al. Salinomycin inhibits osteosarcoma by targeting itstumor stem cells. Cancer Lett. 2011 Dec 1;311(1):113-21.
11. Varelas X, et al. The Hippo pathway regulatesWnt/beta-catenin signaling. Dev Cell. 2010 Apr 20;18(4):579-91.
12. Lu D, et al. Salinomycin inhibits Wnt signaling and selectively induces apoptosis in chronic lymphocyticleukemia cells.Proc Natl Acad Sci U S A. 2011 Aug 9;108(32):13253-7.
13. García-Reyes B, et al. Discovery of Inhibitor of Wnt Production2 (IWP-2) and Related Compounds As Selective ATP-Competitive Inhibitors ofCasein Kinase 1 (CK1) δ/ε. J Med Chem. 2018 May10;61(9):4087-4102.
14. Mathur R, et al. Targeting Wnt pathway in mantle cell lymphoma-initiating cells.J Hematol Oncol. 2015 Jun 6;8:63.
15. Chen B, et al. Small molecule-mediated disruption of Wnt-dependent signaling intissue regeneration and cancer. Nat Chem Biol. 2009 Feb;5(2):100-7.
16. Huang SM, et al. Tankyrase inhibition stabilizes axin andantagonizes Wnt signalling. Nature. 2009 Oct 1;461(7264):614-20
17. Gwak J, et al. Small molecule-based disruption ofthe Axin/β-catenin protein complex regulates mesenchymalstem cell differentiation.Cell Res. 2012 Jan;22(1):237-47.
18. De Sousa E Melo F, et al. Axing Wnt signals. Cell Res. 2012Jan;22(1):9-11.



