
®”(dЁЎng)З°О»ЦГ > КЧн“ > јјРg(shЁҙ)ОДХВ > m5C MeRIP-seqөИҪТКҫm5CРЮп—ФЪ°©°YДНЛҺЦРөДкP(guЁЎn)жIХ{(diЁӨo)ҝШҷCЦЖЦРөД‘Ә(yЁ©ng)УГ
m5C MeRIP-seqөИҪТКҫm5CРЮп—ФЪ°©°YДНЛҺЦРөДкP(guЁЎn)жIХ{(diЁӨo)ҝШҷCЦЖЦРөД‘Ә(yЁ©ng)УГ
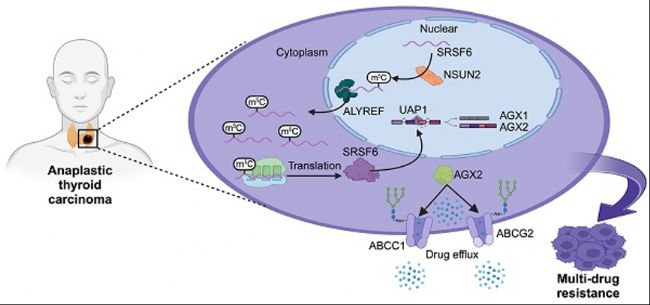
јЧ оПЩОҙ·Ц»Ҝ°©ЈЁAnaplastic Thyroid Cancer, ATCЈ©КЗТ»·NҳOҫЯЗЦТuРФөДјЧ оПЩ°©Ј¬НЁіЈФЪіхФ\•rҫНТСіц¬F(xiЁӨn)ҫЦІҝНнЖЪҪюқҷ»тЯhМҺЮD(zhuЁЈn)ТЖЈ¬еeЯ^БЛЧојСКЦРg(shЁҙ)•rҷCЎЈТтҙЛЈ¬ПөҪy(tЁҜng)»ҜҜҹәН°РПтЦОҜҹҢҰУЪёДЙЖATC»јХЯөДоA(yЁҙ)әуЦБкP(guЁЎn)ЦШТӘЎЈИ»¶шЈ¬ATCҢҰӮчҪy(tЁҜng)ЦОҜҹұн¬F(xiЁӨn)іцп@ЦшөДДНЛҺРФЈ¬Я@К№өГСРҫҝЖдДНЛҺҷCЦЖІў°l(fЁЎ)¬F(xiЁӨn)РВөДЦОҜҹ°РьcіЙһй®”(dЁЎng)„Х(wЁҙ)Ц®јұЎЈҪьДкҒнЈ¬RNAРЮп—ЈЁУИЖдКЗm5CРЮп—Ј©ФЪД[Бц°l(fЁЎ)ЙъәНДНЛҺРФЦРөДЧчУГЦрқuКЬөҪкP(guЁЎn)ЧўЈ¬ө«ЖдФЪATCЦРөДҫЯуwҷCЦЖЙРІ»ЗеіюЎЈ
ҪьИХЈ¬МмҪтбt(yЁ©)ҝЖҙуҢW(xuЁҰ)Д[Бцбt(yЁ©)ФәаҚПтЗ°ҪМКЪҲFк ТФјЧ оПЩОҙ·Ц»Ҝ°©ЈЁATCЈ©һйСРҫҝҢҰПуЈ¬АыУГm5C MeRIP-seqөИјјРg(shЁҙ)ҪТКҫБЛNSUN2ҪйҢ§(dЁЈo)өДm5CРЮп—ФЪATCДНЛҺРФЦРөДЧчУГҷCЦЖЈ¬кUГчБЛNSUN2/SRSF6/UAP1РЕМ–ЭSФЪATC¶аЛҺДНЛҺРФЈЁMultidrug Resistance, MDRЈ©ЦРөДкP(guЁЎn)жIЧчУГЈ¬ІўМбіцНЁЯ^РЎ·ЦЧУNSUN2ТЦЦЖ„©ҝЛ·юДНЛҺРФөДРВІЯВФЎЈЯ@Т»°l(fЁЎ)¬F(xiЁӨn)І»ғHһйATCөДЦОҜҹМṩБЛРВөД°РьcЈ¬ТІһйАнҪвRNAРЮп—ФЪД[БцДНЛҺРФЦРөДЧчУГМṩБЛЦШТӘТА“ю(jЁҙ)ЎЈПакP(guЁЎn)СРҫҝіЙ№ыТФЎ¶NSUN2-mediated m5C modification drives alternative splicing reprogramming and promotes multidrug resistance in anaplastic thyroid cancer through the NSUN2/SRSF6/UAP1 signaling axisЎ·һйо}°l(fЁЎ)ұнУЪЎ¶TheranosticsЎ·лsЦҫЎЈ

ҳЛ(biЁЎo)о}ЈәNSUN2-mediated m5C modification drives alternative splicing reprogramming and promotes multidrug resistance in anaplastic thyroid cancer through the NSUN2/SRSF6/UAP1 signaling axisЈЁNSUN2ҪйҢ§(dЁЈo)өДm5CРЮп—НЁЯ^NSUN2/SRSF6/UAP1РЕМ–ЭSтҢ(qЁұ)„УҝЙЧғјфЗРЦШҫҺіМІўҙЩЯMјЧ оПЩОҙ·Ц»Ҝ°©өД¶аЛҺДНЛҺРФЈ©
°l(fЁЎ)ұн•rйgЈә2025-1-27
°l(fЁЎ)ұнЖЪҝҜЈәTheranostics
У°н‘ТтЧУЈәIF 12.4/Q1
јјРg(shЁҙ)ЖҪЕ_ЈәMeRIP-seqЎўscRNA-seqЎўbulk RNA-seqЈЁТЧ»щТтҪрЕЖјјРg(shЁҙ)Ј©
ұҫСРҫҝНЁЯ^ҢҰATCҳУұҫөДbulkЮD(zhuЁЈn)дӣҪMЈЁbulk RNA-seqЈ©әНҶОјҡ°ыRNAңyРтЈЁscRNA-seqЈ©”ө(shЁҙ)“ю(jЁҙ)ЯMРРИ«Гж·ЦОцЈ¬ТФәYЯxЕc¶аЛҺДНЛҺРФЈЁMDRЈ©ПакP(guЁЎn)өДm5CРЮп—»щТтЎЈлSәуЯMРРIC50ҢҚтһЎўБчКҪјҡ°ыРg(shЁҙ)·ЦОцЈ¬ІўАыУГNsun2»щТтЗГіэөДЧФ°l(fЁЎ)РФјЧ оПЩ°©ЈЁspontaneous tumorigenic ATCЈ©РЎКуДЈРНЈ¬ЧCГчБЛNSUN2ФЪATCЦРҙЩЯMMDRөДЧчУГЎЈһйБЛМҪҫҝNSUN2ҪйҢ§(dЁЈo)өДДНЛҺҷCЦЖЈ¬СРҫҝҳӢ(gЁ°u)ҪЁБЛNSUN2»щТтЗГіэөДATCјҡ°ыПөЈ¬ІўЯMРРЮD(zhuЁЈn)дӣҪMҢW(xuЁҰ)Ўўө°°ЧЩ|(zhЁ¬)ҪMҢW(xuЁҰ)әНMeRIP-seq·ЦОцЎЈҙЛНвЯҖНЁЯ^RNA-seqңyРтәНҝЙЧғјфЗР(alternative splicingЈ¬AS)·ЦОцЈ¬СРҫҝNSUN2ЗГіэәуөДХыуwЧғ»ҜЎЈЯMТ»ІҪНЁЯ^МЗө°°ЧИҫЙ«ЎўЧғРФГвТЯіБөн·әЛШ»ҜЎўәЛЩ|(zhЁ¬)·ЦлxәНPCRөИ·Ҫ·ЁЈ¬МҪУ‘БЛNSUN2/SRSF6/UAP1ЭSөДқ“ФЪҷCЦЖЎЈЧоәуФЪуwНвәНуwғИ(nЁЁi)Фu№АБЛРЎ·ЦЧУNSUN2ТЦЦЖ„©Еcҝ№°©ЛҺОпөД…f(xiЁҰ)Н¬Р§‘Ә(yЁ©ng)ЎЈ
СРҫҝҪY(jiЁҰ)№ыұнГчЈ¬NSUN2ұнЯ_ЕcATCЦРөДMDRп@ЦшПакP(guЁЎn)ЎЈNSUN2Ччһйm5CөД“writer”Ј¬ALYREFЧчһй“reader”Ј¬ЛьӮғ№ІН¬ЧчУГУЪSRSF6 mRNAЈ¬ХTҢ§(dЁЈo)ҝЙЧғјфЗРЦШҫҺіМЈ¬ІўҢўUAP1»щТтөДјфҪУРОКҪҸДAGX1ЮD(zhuЁЈn)ПтAGX2ЎЈҪY(jiЁҰ)№ыұнГчЈ¬AGX2ФцҸҠБЛABCЮD(zhuЁЈn)Я\ө°°ЧөДN-МЗ»щ»ҜЈ¬НЁЯ^ТЦЦЖ·әЛШ»ҜҪйҢ§(dЁЈo)өДҪөҪвТФ·Җ(wЁ§n)¶ЁЯ@Р©ө°°ЧЎЈNSUN2ТЦЦЖ„©НЁЯ^ҪөөНNSUN2Гё»оРФІўңpЙЩПВУО°РҳЛ(biЁЎo)ұнЯ_Ј¬һйҝЛ·юATCЦРөДMDRМṩБЛТ»·NРВөДЎўУРЗ°ҫ°өДЦОҜҹІЯВФЎЈ
ұҫСРҫҝҪТКҫБЛNSUN2/SRSF6/UAP1РЕМ–ЭSФЪATCөДMDRЦРөДкP(guЁЎn)жIЧчУГЈ¬Іўҙ_¶ЁБЛNSUN2Ччһй»ҜҜҹәН°РПтЦОҜҹөД…f(xiЁҰ)Н¬°РьcЎЈ
СРҫҝ·Ҫ·Ё
ЙъОпРЕПўҢW(xuЁҰ)·ЦОцЈәНЁЯ^·ЦОцATCҳУұҫөДbulk RNA-seqңyРт”ө(shЁҙ)“ю(jЁҙ)Ј¬әYЯxЕcMDRПакP(guЁЎn)өДm5CРЮп—»щТтЎЈ
јҡ°ыҢҚтһәН·ЦЧУҷCЦЖСРҫҝЈәАыУГCRISPR/Cas9јјРg(shЁҙ)ҳӢ(gЁ°u)ҪЁNSUN2»щТтЗГіэЈЁknockoutЈ©өДATCјҡ°ыПөЈ¬НЁЯ^RNA-seqЎўө°°ЧЩ|(zhЁ¬)ҪMҢW(xuЁҰ)әНMeRIP-seq·ЦОцөИјјРg(shЁҙ)СРҫҝNSUN2өД№ҰДЬЎЈ
„УОпДЈРНЈәҳӢ(gЁ°u)ҪЁNsun2»щТтЗГіэөДЧФ°l(fЁЎ)РФјЧ оПЩ°©РЎКуДЈРНЈ¬Фu№АNSUN2уwғИ(nЁЁi)ҢҰMDRУ°н‘ЎЈ
ЛҺОпГфёРРФ·ЦОцЈәНЁЯ^IC50ҢҚтһәНБчКҪјҡ°ыРg(shЁҙ)ҷzңyNSUN2ТЦЦЖ„©Еcҝ№°©ЛҺОпөДВ“(liЁўn)әПР§‘Ә(yЁ©ng)ЎЈ
СРҫҝҪY(jiЁҰ)№ы
ЈЁ1Ј©ЙъОпРЕПўҢW(xuЁҰ)·ЦОцҪТКҫATCЦРNSUN2әН¶аЛҺДНЛҺРФЈЁMDRЈ©Ц®йgөДПакP(guЁЎn)РФ
НЁЯ^·ЦОцCancer Therapeutics Response PortalЈЁCTRPЈ©”ө(shЁҙ)“ю(jЁҙ)ҺмЦРөДЛҺОпГфёРРФ”ө(shЁҙ)“ю(jЁҙ)Ј¬°l(fЁЎ)¬F(xiЁӨn)NSUN2өДұнЯ_Еc¶а·Nҝ№°©ЛҺОпЈЁ°ьАЁ»ҜҜҹЛҺОпәНАТ°ұЛбјӨГёТЦЦЖ„©Ј©өДIC50ЦөіКХэПакP(guЁЎn)ЎЈ
АыУГGEO”ө(shЁҙ)“ю(jЁҙ)ҺмЦРөДATCЮD(zhuЁЈn)дӣҪM”ө(shЁҙ)“ю(jЁҙ)Ј¬Ңў»јХЯ·ЦһйёЯNSUN2ұнЯ_ҪMәНөНNSUN2ұнЯ_ҪMЈ¬°l(fЁЎ)¬F(xiЁӨn)ёЯNSUN2ұнЯ_Еcјҡ°ы·ЦБСЎўУРҪz·ЦБСЎўјҡ°ыЦЬЖЪөИ»ҜҜҹ°РПтЯ^іМПакP(guЁЎn)ЎЈ
scRNA-seqңyРт·ЦОцп@КҫЈ¬NSUN2ФЪATCјҡ°ыЦРөДұнЯ_Еc¶а·N»ҜҜҹЛҺОпәНTKIөДДНЛҺРФПакP(guЁЎn)ЎЈ
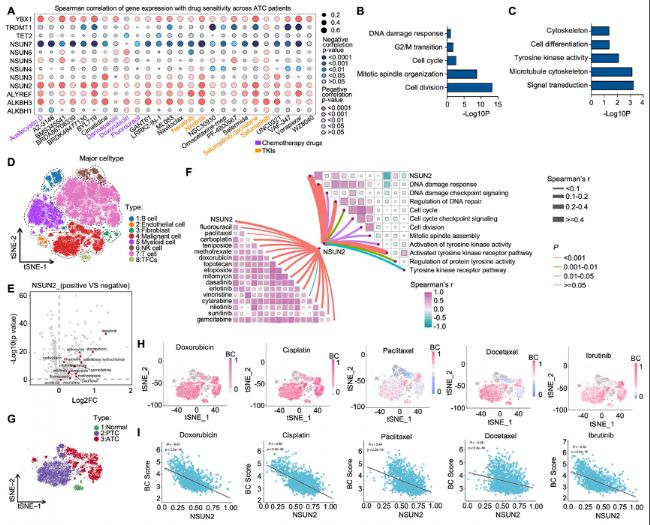
ҲD1ЈәЙъОпРЕПўҢW(xuЁҰ)·ЦОцҪТКҫNSUN2ЕcATCЦР¶аЛҺДНЛҺРФЈЁMDRЈ©өДПакP(guЁЎn)РФЎЈ
AЈ©ҒнЧФCTRP”ө(shЁҙ)“ю(jЁҙ)ҺмөДATC»јХЯ»щТтұнЯ_ЕcЛҺОпГфёРРФөДSpearmanПакP(guЁЎn)РФ·ЦОцЎЈ
B-CЈ©ёЯNSUN2ұнЯ_ҪMЕcөНNSUN2ұнЯ_ҪMЦ®йgІо®җұнЯ_»щТтЈЁDEGsЈ©өДё»јҜНЁВ·ҝӮҪY(jiЁҰ)ЎЈ
DЈ©ATCөДscRNA-seq”ө(shЁҙ)“ю(jЁҙ)өДtSNEҲDЈ¬Х№КҫБЛиb¶ЁіцөДјҡ°ыclustersЈЁn=10Ј©ЎЈ
EЈ©ЙўьcҲDп@КҫNSUN2к–РФјҡ°ыЕcNSUN2кҺРФјҡ°ыөДlog2ұ¶Чғ»ҜЈЁlog2FCЈ©ј°ЖдЕcГҝ·NЛҺОпоA(yЁҙ)ңyөДIC50ЗъҫҖПВГж·eЈЁAUCЈ©ЦөөДПакP(guЁЎn)РФЎЈ
FЈ©әыөыҲDХ№КҫNSUN2ұнЯ_ЎўЛҺОпГфёРРФЕc»щТтұҫуwЈЁGOЈ©НЁВ··ЦОцЦ®йgөДПакP(guЁЎn)РФЎЈ
GЈ©ATCөДscRNA-seq”ө(shЁҙ)“ю(jЁҙ)өДtSNEҲDЈ¬Х№КҫБЛиb¶ЁіцөДјҡ°ыclustersЈЁn=10Ј©ЎЈ
HЈ©К№УГBeyondcellҢҰGEO”ө(shЁҙ)“ю(jЁҙ)јҜҳУұҫЯMРРөДҶОјҡ°ыЛҺОпГфёРРФ·ЦОцЎЈ
IЈ©NSUN2ұнЯ_ЕcBeyondcellФu·ЦЦ®йgөДПакP(guЁЎn)РФЎЈ
ЈЁ2Ј©ATCЦРNSUN2ХTҢ§(dЁЈo)өДMDRИЎӣQУЪЖдјЧ»щЮD(zhuЁЈn)ТЖГё»оРФ
ФЪATCјҡ°ыПөЦРЗГөН»тЗГіэNSUN2әуЈ¬јҡ°ыҢҰ¶а·NЛҺОпөДГфёРРФп@ЦшФцјУЈ¬ұнГчNSUN2ұнЯ_ЕcMDRГЬЗРПакP(guЁЎn)ЎЈ
НЁЯ^ҳӢ(gЁ°u)ҪЁNSUN2ҙЯ»Ҝ»оРФН»ЧғуwЈЁC271AЈ©Ј¬°l(fЁЎ)¬F(xiЁӨn)Ц»УРТ°ЙъРНNSUN2ДЬүт»ЦҸН(fЁҙ)m5CРЮп—»оРФәНЛҺОпДНКЬРФЈ¬ұнГчNSUN2өДјЧ»щЮD(zhuЁЈn)ТЖГё»оРФКЗЖдХTҢ§(dЁЈo)MDRөДкP(guЁЎn)жIЎЈ
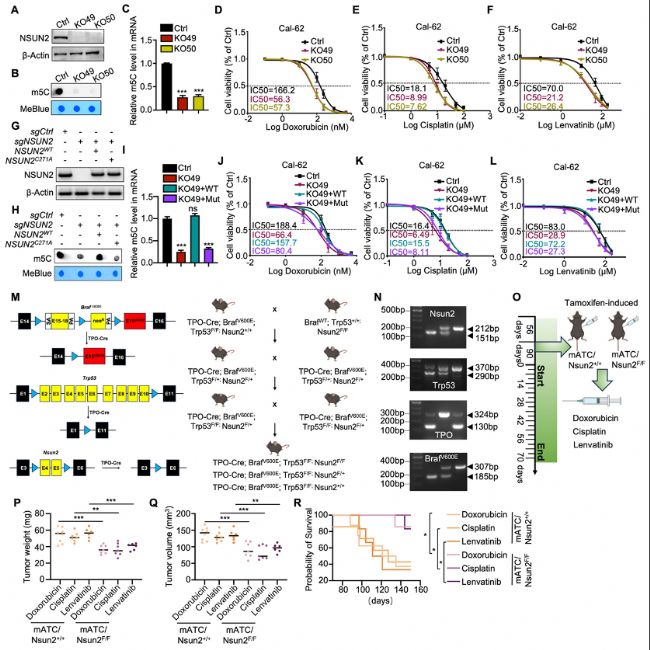
BЈ©НЁЯ^ьcлsҪ»ҢҚтһҷzңyNSUN2»щТтЗГіэҢҰCal-62јҡ°ыmRNAЮD(zhuЁЈn)дӣҪMЦРm5CЛ®ЖҪөДУ°н‘ЎЈ
CЈ©НЁЯ^ұИЙ«·Ёm5C¶ЁБҝҢҚтһҙ_ХJNSUN2»щТтЗГіэәуCal-62јҡ°ыЦРm5CЛ®ЖҪөДЧғ»ҜЎЈ
D-FЈ©НЁЯ^јҡ°ыЙъйLТЦЦЖҢҚтһФu№АNSUN2»щТтЗГіэҢҰCal-62јҡ°ыҢҰ¶аИбұИРЗЈЁdoxorubicinЈ©ЎўнҳгKЈЁcisplatinЈ©әНҒц·ҘМжДбЈЁlenvatinibЈ©ГфёРРФөДУ°н‘ЎЈ
GЈ©НЁЯ^Western blot·ЦОцҷzңyCal-62јҡ°ыҪӣ(jЁ©ng)І»Н¬МҺАнәуNSUN2ө°°ЧөДПаҢҰұнЯ_БҝЎЈ
HЈ©НЁЯ^ьcлsҪ»ҢҚтһҷzңyҪӣ(jЁ©ng)ЭdуwМҺАнәуCal-62јҡ°ыmRNAЮD(zhuЁЈn)дӣҪMЦРm5CШS¶ИЎЈ
IЈ©НЁЯ^ұИЙ«·Ёm5C¶ЁБҝҢҚтһҙ_ХJҪӣ(jЁ©ng)МҺАнәуCal-62јҡ°ыЦРm5CЛ®ЖҪЧғ»ҜЎЈ
J-LЈ©НЁЯ^јҡ°ыЙъйLТЦЦЖҢҚтһФu№АҪӣ(jЁ©ng)І»Н¬ЭdуwЮD(zhuЁЈn)ИҫөДCal-62јҡ°ыөДIC50ЦөЎЈ
MЈ©ЧФ°l(fЁЎ)РФATCРЎКуДЈРНөДҳӢ(gЁ°u)ҪЁ·Ҫ°ёЈ¬°ьАЁ°РПтІЯВФЈЁЧуЈ©әН·ұЦі·Ҫ°ёЈЁУТЈ©ЎЈ
NЈ©РЎКу»щТт·ЦРНҪY(jiЁҰ)№ыЈ¬ЖдЦРTPO-creөДDNAДэДz—lҺ§ҙуРЎһй130/324bpЈ¬BrafV600Eһй185/307bpЈ¬Trp53flox/floxһй290/370bpЈ¬Nsun2flox/floxһй151/212bpЎЈ
OЈ©ФЪ»щТт№ӨіМРЎКуЦР¶аИбұИРЗЎўнҳгKәНҒц·ҘМжДбөДҪoЛҺ·Ҫ°ёЎЈmATCЈЁРЎКујЧ оПЩОҙ·Ц»Ҝ°©Ј©ЎЈ
PЈ©ЙўьcҲDп@КҫёчҪMРЎКуөДЧоҪKД[БцЦШБҝЎЈ
QЈ©ЙўьcҲDп@КҫёчҪMРЎКуөДЧоҪKД[Бцуw·eЎЈ
RЈ©п@КҫёчҪMATCРЎКуөДЙъҙжЗъҫҖЎЈ
ЈЁ3Ј©ФЪATCЦРЈ¬NSUN2ҪйҢ§(dЁЈo)өДm5CРЮп—НЁЯ^°РПтSRSF6тҢ(qЁұ)„УҝЙЧғјфЗРЦШҫҺіМ
ө°°ЧЩ|(zhЁ¬)ҪMҢW(xuЁҰ)·ЦОцп@КҫЈ¬NSUN2ЗГіэәуЈ¬ЕcRNAјфҪУПакP(guЁЎn)өДө°°ЧұнЯ_°l(fЁЎ)Йъп@ЦшЧғ»ҜЈ¬ЖдЦРSRSF6ө°°ЧұнЯ_п@ЦшПВХ{(diЁӨo)Ј¬¶шЖдmRNAЛ®ЖҪІ»ЧғЎЈ
MeRIP-seqәНqPCRҪY(jiЁҰ)№ыұнГчЈ¬NSUN2НЁЯ^m5CРЮп—Х{(diЁӨo)№қ(jiЁҰ)SRSF6 mRNAөДәЛЩ|(zhЁ¬)ЮD(zhuЁЈn)Я\Ј¬ЯM¶шУ°н‘Ждө°°ЧұнЯ_ЎЈ
НЁЯ^Н»ЧғSRSF6 mRNAЙПөДm5CРЮп—О»ьcЈ¬°l(fЁЎ)¬F(xiЁӨn)Я@Р©О»ьcҢҰSRSF6өДәЛЩ|(zhЁ¬)ЮD(zhuЁЈn)Я\ЦБкP(guЁЎn)ЦШТӘЎЈ

BЈ©Cal-62јҡ°ыЦРNSUN2»щТтЗГіэәуTMT-MS”ө(shЁҙ)“ю(jЁҙ)өДGOё»јҜ·ЦОцЎЈ
CЈ©Cal-62јҡ°ыЦРNSUN2»щТтЗГіэәуІо®җјфҪУ»щТтөД°Щ·ЦұИјфҪУ°ьә¬ЈЁΔPSIЈ©өД»рЙҪҲDЎЈ
DЈ©Cal-62јҡ°ыЦРNSUN2»щТтЗГіэәуп@ЦшΔPSIЧғ»ҜөДРЎМбЗЩҲDЎЈ
EЈ©Cal-62јҡ°ыЦРNSUN2»щТтЗГіэәуSF3A/BЎўU2AFәЛРДҸН(fЁҙ)әПуwәНhnRNPјТЧеЦРDEGҹбҲDЎЈ
FЈ©Cal-62јҡ°ыЦРNSUN2»щТтЗГіэәуTMT-MS”ө(shЁҙ)“ю(jЁҙ)ЦРSRSF6ө°°ЧЛ®ЖҪөДҪy(tЁҜng)УӢ·ЦОцЎЈ
GЈ©Cal-62јҡ°ыЦРNSUN2»щТтЗГіэәуЦёКҫө°°ЧөДWestern blot·ЦОцЎЈ
HЈ©Cal-62јҡ°ыЦРNSUN2»щТтЗГіэәуSRSF6»щТтЧщЦЬҮъөДMeRIP-seq”ө(shЁҙ)“ю(jЁҙ)өДIGVЬүЫEҲDЎЈ
IЈ©Cal-62јҡ°ыЦРNSUN2»щТтЗГіэәуSRSF6 mRNAЦРm5Cё»јҜөДMeRIP-qPCRҷzңyЎЈ
JЈ©Ҫӣ(jЁ©ng)ЦёКҫЭdуwМҺАнөДCal-62јҡ°ыЦРSRSF6 mRNAЦРm5Cё»јҜөДMeRIP-qPCR·ЦОцЎЈ
KЈ©NSUN2»щТтЗГіэәуCal-62јҡ°ыЦРSRSF6 mRNAөДәЛЩ|(zhЁ¬)·ЦІјqRT-PCR·ЦОцЎЈ
LЈ©Ҫӣ(jЁ©ng)ЦёКҫЭdуwМҺАнөДCal-62јҡ°ыЦРSRSF6 mRNAөДәЛЩ|(zhЁ¬)·ЦІјЎЈ
MЈ©Ҫӣ(jЁ©ng)ЦёКҫЭdуwМҺАнөДCal-62јҡ°ыЦРЦёКҫө°°ЧөДWestern blot·ЦОцЎЈ
NЈ©SRSF6ЦРөДөдРНm5CҪY(jiЁҰ)әПО»ьcЎЈн”Іҝһйm5CҪY(jiЁҰ)әПО»ьcөДmotifРтБРЈ»өЧІҝһйSRSF6ЦРиb¶ЁөДm5CО»ьcЎЈ
OЈ©Ҫӣ(jЁ©ng)ЦёКҫЭdуwМҺАнөДCal-62јҡ°ыЦРSRSF6 mRNAЦРm5Cё»јҜөДMeRIP-qPCR¶ЁБҝЎЈ
PЈ©Ҫӣ(jЁ©ng)ЦёКҫЭdуwМҺАнөДCal-62јҡ°ыЦРSRSF6 mRNAөДәЛЩ|(zhЁ¬)·ЦІјЎЈ
ЈЁ4Ј©NSUN2НЁЯ^ФцҸҠABCЮD(zhuЁЈn)Я\ө°°ЧөДN-МЗ»щ»ҜҙЩЯMATCЦРөДMDR
ЮD(zhuЁЈn)дӣҪM·ЦОцәНKEGGНЁВ··ЦОцп@КҫЈ¬NSUN2…ўЕcN-МЗжңЙъОпәПіЙЯ^іМЎЈ
НЁЯ^МЗө°°ЧИҫЙ«әНө°°ЧЩ|(zhЁ¬)УЎЫE·ЦОцЈ¬°l(fЁЎ)¬F(xiЁӨn)NSUN2ЗГіэәуЈ¬ABCЮD(zhuЁЈn)Я\ө°°ЧЈЁABCC1әНABCG2Ј©өДN-МЗ»щ»ҜЛ®ЖҪҪөөНЈ¬ө°°Ч·Җ(wЁ§n)¶ЁРФПВҪөЎЈ
БчКҪјҡ°ыРg(shЁҙ)ҷzңyп@КҫЈ¬NSUN2ЗГіэјҡ°ыҢҰЛҺОп”zИЎФцјУЈ¬ұнГчЛҺОпНвЕЕңpЙЩЈ¬ДНЛҺРФҪөөНЎЈ
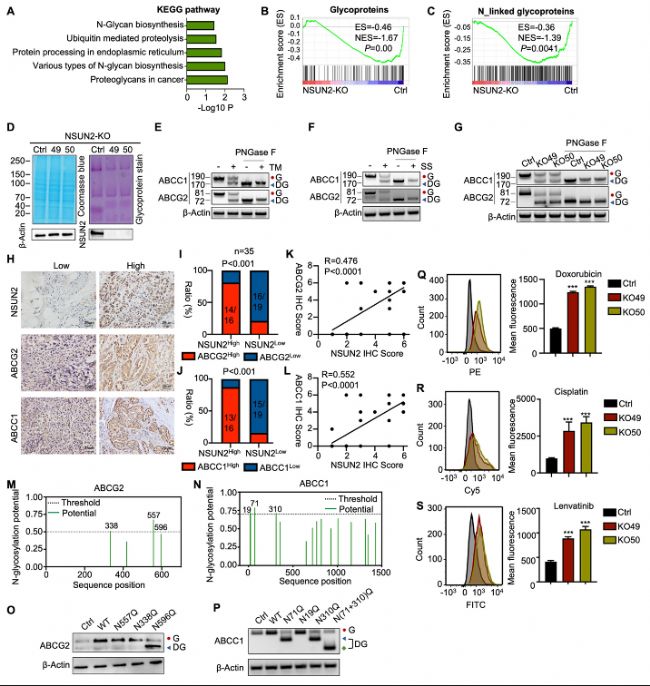
B-CЈ©Cal-62јҡ°ыЦРNSUN2»щТтЗГіэәуЮD(zhuЁЈn)дӣ”ө(shЁҙ)“ю(jЁҙ)өД»щТтјҜё»јҜ·ЦОцЈЁGSEAЈ©ЎЈ
DЈ©ҝӮө°°ЧөДҝјсRЛ№ББЛ{ИҫЙ«ЈЁЧуЈ©Ј¬NSUN2»щТтЗГіэәуCal-62јҡ°ыөДМЗө°°ЧИҫЙ«ҪY(jiЁҰ)№ыЈЁУТЈ©ЎЈ
E-FЈ©Ҫӣ(jЁ©ng)»тОҙҪӣ(jЁ©ng)TMЈЁtunicamycinЈ©ЎўSSЈЁswainsonineЈ©МҺАнөДCal-62јҡ°ыЦРЦёКҫө°°ЧЛ®ЖҪөДWestern blot·ЦОцЎЈјҡ°ыБСҪвҳУұҫҪӣ(jЁ©ng)»тОҙҪӣ(jЁ©ng)PNGase FМҺАнЎЈ“G”ұнКҫN-МЗ»щ»ҜЈ¬“DG”ұнКҫN-МЗ»щ»ҜңpЙЩЎЈ
GЈ©NSUN2»щТтЗГіэөДCal-62јҡ°ыЦРЦёКҫө°°ЧЛ®ЖҪөДWestern blot·ЦОцЎЈ
HЈ©ҙъұнРФГвТЯҪM»ҜЈЁIHCЈ©ҲDПсп@Кҫ35АэATCҪMҝ—ЦРNSUN2ЎўABCG2әНABCC1өДұнЯ_ЗйӣrЎЈ
I-JЈ©ІЙУГҝЁ·Ҫҷzтһ·ЦОцNSUN2ЕcABCG2»тABCC1ұнЯ_Ц®йgөДПакP(guЁЎn)РФЎЈ
K-LЈ©NSUN2ЕcABCG2»тABCC1өДIHCФu·ЦөДПакP(guЁЎn)РФ·ЦОцЎЈ
M-NЈ©К№УГNetNglyc·ю„Х(wЁҙ)ЖчоA(yЁҙ)ңyөДABCG2әНABCC1өДқ“ФЪN-МЗ»щ»ҜМ춬хЈ°·О»ьcЎЈ
O-PЈ©·Ц„eФЪЮD(zhuЁЈn)ИҫБЛN557QЎўN388QЎўN596Q»тN71QЎўN19QЎўN310QЎўN(71+310)QН»ЧғЩ|(zhЁ¬)БЈөДCal-62јҡ°ыЦРЈ¬НЁЯ^Western blotҷzңyABCG2әНABCC1өДұнЯ_ЗйӣrЎЈ
Q-SЈ©NSUN2»щТтЗГіэәуCal-62јҡ°ыЦР¶аИбұИРЗЎўнҳгKәНҒц·ҘМжДбөДјҡ°ығИ(nЁЁi)·eАЫЗйӣrөДБчКҪјҡ°ығx·ЦОцЎЈУТӮИ(cЁЁ)п@КҫЖҪҫщҹЙ№вҸҠ¶ИөД¶ЁБҝҪY(jiЁҰ)№ыЎЈ
ЈЁ5Ј©NSUN2НЁЯ^N-МЗ»щ»ҜФцҸҠABCЮD(zhuЁЈn)Я\ө°°ЧөД·Җ(wЁ§n)¶ЁРФ
НЁЯ^western blot ·ЦОцәН·әЛШ»ҜҢҚтһЈ¬°l(fЁЎ)¬F(xiЁӨn)NSUN2ЗГіэәуЈ¬ABCC1әНABCG2өД·әЛШ»ҜЛ®ЖҪФцјУЈ¬ұнГчN-МЗ»щ»ҜИұК§Ң§(dЁЈo)ЦВЯ@Р©ө°°ЧёьИЭТЧұ»ҪөҪвЎЈ
К№УГө°°ЧГёуwТЦЦЖ„©MG132ҝЙТФІҝ·Ц»ЦҸН(fЁҙ)ABCC1әНABCG2өДө°°ЧЛ®ЖҪЈ¬ЯMТ»ІҪЧCҢҚБЛN-МЗ»щ»ҜҢҰө°°Ч·Җ(wЁ§n)¶ЁРФөДУ°н‘ЎЈ
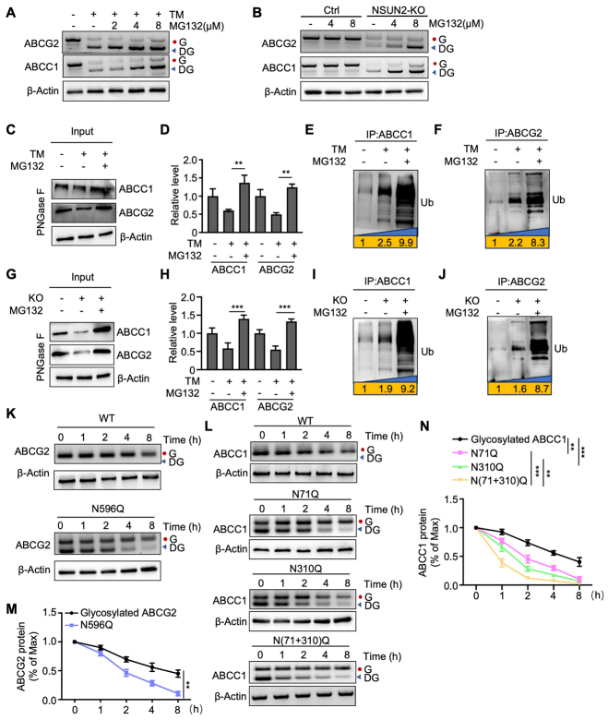
BЈ©NSUN2»щТтЗГіэөДCal-62јҡ°ыҪӣ(jЁ©ng)І»Н¬қв¶ИөДMG132МҺАнәуЈ¬К№УГЦё¶Ёҝ№уwЯMРРWestern blot·ЦОцЎЈ
CЈ©Ҫӣ(jЁ©ng)TMәНMG132МҺАнөДCal-62јҡ°ыЦРЦёКҫө°°ЧЛ®ЖҪөДWestern blot·ЦОцЎЈ
DЈ©К№УГImageJЬӣјюҢҰABCC1әНABCG2ө°°ЧЛ®ЖҪЯMРР¶ЁБҝ·ЦОцЎЈ
E-FЈ©Ҫӣ(jЁ©ng)TMәНMG132МҺАнөДCal-62јҡ°ыЯMРРABCC1»тABCG2ГвТЯіБөнЈЁIPЈ©Ј¬лSәуК№УГҝ№·әЛШҝ№уwЯMРРWestern blot·ЦОцЎЈ
GЈ©NSUN2»щТтЗГіэЗТҪӣ(jЁ©ng)MG132МҺАнөДCal-62јҡ°ыЦРЦёКҫө°°ЧЛ®ЖҪөДWestern blot·ЦОцЎЈ
HЈ©К№УГImageJЬӣјюҢҰABCC1әНABCG2ө°°ЧҸҠ¶ИЯMРР¶ЁБҝ·ЦОцЎЈ
I-JЈ©NSUN2»щТтЗГіэЗТҪӣ(jЁ©ng)MG132МҺАнөДCal-62јҡ°ыЯMРРABCC1»тABCG2ГвТЯіБөнЈЁIPЈ©Ј¬лSәуК№УГҝ№·әЛШҝ№уwЯMРРWestern blot·ЦОцЎЈ
K-LЈ©ФЪCal-62јҡ°ыЦРұнЯ_өДТ°ЙъРНЈЁWTЈ©»тЦёКҫөДABCG2/ABCC1Н»ЧғуwҪӣ(jЁ©ng)125μg/mLӯh(huЁўn)јәхЈҒҶ°·ЈЁCHXЈ©МҺАнМШ¶Ё•rйgйgёфәуЈ¬НЁЯ^Western blotЯMРР·ЦОцЎЈ
M-NЈ©К№УГImageJЬӣјюҢҰABCG2»тABCC1өДө°°ЧҸҠ¶И¶ЁБҝ·ЦОцЈ¬ІўТФβ-ActinЛ®ЖҪҡwТ»»ҜЎЈ
ЈЁ6Ј©NSUN2НЁЯ^ҪйҢ§(dЁЈo)UAP1өҪSRSF6өДҝЙЧғјфЗРҒнҙЩЯMN-МЗ»щ»Ҝ
UAP1КЗN-МЗ»щ»Ҝ·ҙ‘Ә(yЁ©ng)ЦРUDP-GlcNAcәПіЙөДкP(guЁЎn)жIГёЈ¬ЖдУРғЙ·NҒҶРНЈЁAGX1әНAGX2Ј©ЎЈ
NSUN2ЗГіэәуЈ¬UAP1өДјфҪУРОКҪҸДAGX2ЮD(zhuЁЈn)Чғ?yЁӯu)йAGX1Ј¬Ң§(dЁЈo)ЦВN-МЗ»щ»Ҝ»оРФҪөөНЎЈ
НЁЯ^Я^ұнЯ_SRSF6Ј¬ҝЙТФДжЮD(zhuЁЈn)Я@Т»јфҪУЧғ»ҜЈ¬»ЦҸН(fЁҙ)N-МЗ»щ»ҜЛ®ЖҪәНЛҺОпДНКЬРФЎЈ

BЈ©SashimiҲDп@КҫNSUN2»щТтЗГіэәуCal-62јҡ°ыЦРUAP1јфЗРКВјюөДЧғ»ҜЎЈ
CЈ©ҢҰХХҪMЕcNSUN2»щТтЗГіэөДCal-62јҡ°ыЦРUAP1јфЗРөДRT-PCR·ЦОцЎЈ
DЈ©ЧФ°l(fЁЎ)РФРЎКуATCД[БцЦРUAP1өДјфЗРКВјюөДRT-PCRЈ¬УР»тӣ]УРNsun2»щТтЗГіэЎЈ
EЈ©Ҫӣ(jЁ©ng)ЦёКҫЭdуwМҺАнөДCal-62јҡ°ыЦРUAP1јфЗРКВјюөДRT-PCRЈЁЙПҲDЈ©ЎЈWestern blot·ЦОцБЛSRSF6ЦШҪЁЈЁПВҲDЈ©ЎЈ
FЈ©ҷzңyҝӮө°°ЧөДҝјсRЛ№ББЛ{ИҫЙ«ЈЁЧуЈ©ЎЈҪӣ(jЁ©ng)ЦёКҫЭdуwМҺАнөДCal-62јҡ°ыМЗө°°ЧИҫЙ«ЈЁУТЈ©ЎЈ
GЈ©Ҫӣ(jЁ©ng)ЦёКҫЭdуwМҺАнөДCal-62јҡ°ыЦРЦёКҫө°°ЧЛ®ЖҪөДWestern blot·ЦОцЎЈ
H-JЈ©Ҫӣ(jЁ©ng)ЦёКҫЭdуwМҺАнөДCal-62јҡ°ыЦР¶аИбұИРЗЎўнҳгKәНҒц·ҘМжДбөДјҡ°ығИ(nЁЁi)·eАЫЗйӣrөДБчКҪјҡ°ығx·ЦОцЎЈУТӮИ(cЁЁ)п@КҫБЛЖҪҫщҹЙ№вҸҠ¶ИөД¶ЁБҝҪY(jiЁҰ)№ыЎЈ
K-MЈ©ЦёКҫЭdуwМҺАнCal-62јҡ°ыҢҰ¶аИбұИРЗЎўнҳгKәНҒц·ҘМжДбГфёРУ°н‘јҡ°ыЙъйLТЦЦЖҢҚтһЎЈ
N-PЈ©ЦёКҫЭdуwМҺАнCal-62јҡ°ыҢҰ¶аИбұИРЗЎўнҳгKәНҒц·ҘМжДб·ҙ‘Ә(yЁ©ng)У°н‘өДД[БцЙъйLЗъҫҖЎЈ
Q-SЈ©ЦёКҫЭdуwМҺАнCal-62јҡ°ыҢҰ¶аИбұИРЗЎўнҳгKәНҒц·ҘМжДб·ҙ‘Ә(yЁ©ng)У°н‘Д[БцЧоҪKЦШБҝЎЈ
ЈЁ7Ј©Ччһйm5CөД“reader”Ј¬ ALYREFҝЙТФиb¶ЁSRSF6ІўҢўЖдҸДјҡ°ыәЛЮD(zhuЁЈn)Я\ЦБјҡ°ыЩ|(zhЁ¬)
НЁЯ^RNAГвТЯіБөнЈЁRIPЈ©әНRNAПВАӯҢҚтһЈ¬°l(fЁЎ)¬F(xiЁӨn)ALYREFДЬүтЦұҪУҪY(jiЁҰ)әПm5CРЮп—өДSRSF6 mRNAЈ¬ІўҢўЖдҸДјҡ°ыәЛЮD(zhuЁЈn)Я\ЦБјҡ°ыЩ|(zhЁ¬)ЎЈ
ALYREFөДK171AН»Чғуwп@ЦшҪөөНҢҰm5C -SRSF6ҪY(jiЁҰ)әПДЬБҰЈ¬ұнГчALYREFКЗm5CРЮп—өД“reader”ЎЈ
ALYREFЗГөН»тЯ^ұнЯ_ҝЙТФХ{(diЁӨo)ҝШSRSF6өДјҡ°ыәЛЩ|(zhЁ¬)ЮD(zhuЁЈn)Я\әНө°°ЧұнЯ_Ј¬ЯM¶шУ°н‘ЛҺОпДНКЬРФЎЈ
CЈ©ҹбҲDп@КҫФЪNSUN2»щТтЗГіэ»тALYREFЎўYBX1әНSRSF2»щТтЗГөНөДCal-62јҡ°ыЦРЕc¶аИбұИРЗәНнҳгKДНЛҺРФПакP(guЁЎn)өД»щТтЎЈ
DЈ©НЁЯ^RIP-qPCRҷzңyALYREFЕcCal-62јҡ°ыЦРSRSF6өДҪY(jiЁҰ)әПЎЈ
EЈ©НЁЯ^Western blot·ЦОцҸДSRSF6ЈЁm5CЈ©ПВАӯҢҚтһЦР«@өГөДЦёКҫө°°ЧЎЈ
FЈ©НЁЯ^RIP-qPCRұИЭ^ФЪҪӣ(jЁ©ng)ЦёКҫЭdуwМҺАнөДCal-62јҡ°ыЦРЈ¬Т°ЙъРНALYREFЈЁALYREF-WTЈ©әНK171AН»ЧғРНALYREFЈЁALYREF-K171AЈ©ЕcSRSF6өДҪY(jiЁҰ)әПЎЈ
GЈ©НЁЯ^RIP-qPCRФu№АNSUN2»щТтЗГіэәуCal-62јҡ°ыЦРALYREFЕcSRSF6 mRNAөДҪY(jiЁҰ)әПЎЈ
HЈ©НЁЯ^RIP-qPCRФu№АҪӣ(jЁ©ng)ЦёКҫЭdуwМҺАнөДCal-62јҡ°ыЦРALYREFЕcSRSF6 mRNAөДҪY(jiЁҰ)әПЎЈ
IЈ©НЁЯ^qRT-PCRҢҚтһФu№АALYREF»щТтЗГөНөДCal-62јҡ°ыЦРSRSF6 mRNAөДәЛЩ|(zhЁ¬)·ЦІјЎЈ
JЈ©ФЪЯ^ұнЯ_ALYREF»тALYREF-K171AН»ЧғуwөДCal-62јҡ°ыЦРЈ¬SRSF6 mRNAөДәЛЩ|(zhЁ¬)·ЦІјЎЈ
KЈ©Ҫӣ(jЁ©ng)ЦёКҫЭdуwМҺАнөДCal-62јҡ°ыЦРSRSF6 mRNAөДәЛЩ|(zhЁ¬)·ЦІјЎЈ
L-NЈ©јҡ°ыЙъйLТЦЦЖҢҚтһФu№АЦёКҫЭdуwМҺАнҢҰCal-62јҡ°ыҢҰ¶аИбұИРЗЎўнҳгKәНҒц·ҘМжДб·ҙ‘Ә(yЁ©ng)өДУ°н‘ЎЈ
OЈ©НЁЯ^Western blot·ЦОцҪӣ(jЁ©ng)ЦёКҫЭdуwМҺАнөДCal-62јҡ°ыЦРЦёКҫө°°ЧөДЛ®ЖҪЎЈ
PЈ©NSUN2»щТтЗГіэөДCal-62јҡ°ыЦРALYREFөДГвТЯҹЙ№вИҫЙ«ЈЁЧуӮИ(cЁЁ)Ј©ТФј°әЛ/Щ|(zhЁ¬)ALYREF·ЦІјөД¶ЁБҝ·ЦОцЈЁУТӮИ(cЁЁ)Ј©ЎЈ
QЈ©НЁЯ^Western blot·ЦОцЈЁЧуӮИ(cЁЁ)Ј©ТФј°Па‘Ә(yЁ©ng)өД¶ЁБҝ·ЦОцЈЁУТӮИ(cЁЁ)Ј©Ј¬Х№КҫҢҰХХҪMәНNSUN2»щТтЗГіэөДCal-62јҡ°ыЦРәЛәНЩ|(zhЁ¬)ALYREFөДЛ®ЖҪЈ¬PARP1әНTUBULIN·Ц„eЧчһйәЛәНЩ|(zhЁ¬)ҳЛ(biЁЎo)УӣЎЈ
ЈЁ8Ј©РЎ·ЦЧУNSUN2ТЦЦЖ„©ФцҸҠATCЦР¶аЛҺГфёРРФ
ФO(shЁЁ)УӢІўәПіЙРЎ·ЦЧУNSUN2ТЦЦЖ„©ЈЁNSUN2iЈ©Ј¬ДЬүтп@ЦшҪөөНATCјҡ°ыЦРөДm5CЛ®ЖҪЎЈ
NSUN2iМҺАнәуЈ¬SRSF6өДәЛЩ|(zhЁ¬)ЮD(zhuЁЈn)Я\КЬЧиЈ¬UAP1јфҪУРОКҪёДЧғЈ¬N-МЗ»щ»ҜЛ®ЖҪҪөөНЈ¬ABCЮD(zhuЁЈn)Я\ө°°Ч·Җ(wЁ§n)¶ЁРФПВҪөЎЈ
уwНвҢҚтһәН„УОпДЈРНҢҚтһҫщп@КҫЈ¬NSUN2iЕc»ҜҜҹЛҺОп»тTKIВ“(liЁўn)әПК№УГҝЙТФп@ЦшФцҸҠЛҺОпөДҝ№Д[БцР§№ыЎЈ
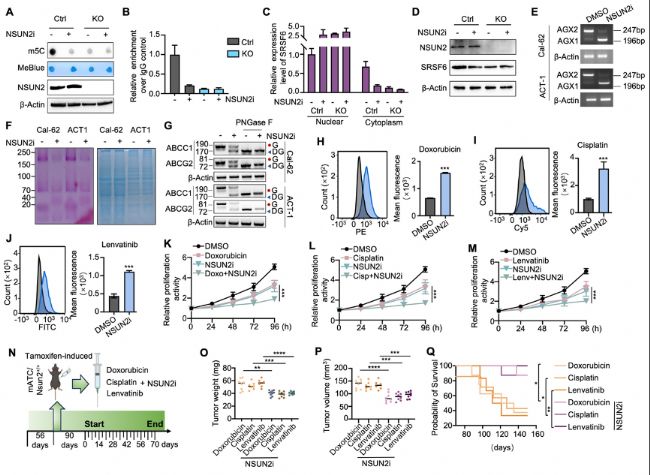
BЈ©»щУЪRT-qPCRөДMeRIPҷzңyNSUN2»щТтЗГіэөДҪӣ(jЁ©ng)»тОҙҪӣ(jЁ©ng)NSUN2ТЦЦЖ„©МҺАнөДCal-62јҡ°ыЦРSRSF6 mRNAөДm5Cё»јҜ·ЦОцЎЈ
CЈ©НЁЯ^qRT-PCRҢҚтһФu№АNSUN2»щТтЗГіэөДҪӣ(jЁ©ng)»тОҙҪӣ(jЁ©ng)NSUN2ТЦЦЖ„©МҺАнөДCal-62јҡ°ыЦРSRSF6 mRNAөДәЛЩ|(zhЁ¬)·ЦІјЎЈ
DЈ©НЁЯ^Western blot·ЦОцNSUN2»щТтЗГіэөДҪӣ(jЁ©ng)»тОҙҪӣ(jЁ©ng)NSUN2ТЦЦЖ„©МҺАнөДCal-62јҡ°ыЦРЦёКҫө°°ЧөДЛ®ЖҪЎЈ
EЈ©RT-PCRп@КҫҪӣ(jЁ©ng)»тОҙҪӣ(jЁ©ng)NSUN2ТЦЦЖ„©МҺАнөДCal-62јҡ°ыәНACT-1јҡ°ыЦРUAP1өДјфҪУКВјюЎЈ
FЈ©ҷzңyҝӮө°°ЧөДҝјсRЛ№ББЛ{ИҫЙ«ЈЁЧуЈ©Ј¬Ҫӣ(jЁ©ng)»тОҙҪӣ(jЁ©ng)NSUN2ТЦЦЖ„©МҺАнөДCal-62јҡ°ыәНACT-1јҡ°ыөДМЗө°°ЧИҫЙ«ЈЁУТЈ©ЎЈ
GЈ©НЁЯ^Western blot·ЦОцҪӣ(jЁ©ng)»тОҙҪӣ(jЁ©ng)NSUN2ТЦЦЖ„©МҺАнөДCal-62јҡ°ыәНACT-1јҡ°ыЦРЦёКҫө°°ЧөДЛ®ЖҪЎЈ
H-JЈ©НЁЯ^БчКҪјҡ°ығx·ЦОцҪӣ(jЁ©ng)»тОҙҪӣ(jЁ©ng)NSUN2ТЦЦЖ„©МҺАнөДCal-62јҡ°ыЦР¶аИбұИРЗЎўнҳгKәНҒц·ҘМжДбөДјҡ°ығИ(nЁЁi)·eАЫЗйӣrЎЈУТӮИ(cЁЁ)п@КҫЖҪҫщҹЙ№вҸҠ¶ИөД¶ЁБҝҪY(jiЁҰ)№ыЎЈ
K-MЈ©јҡ°ыЙъйLТЦЦЖҢҚтһФu№АNSUN2ТЦЦЖ„©Еc¶аИбұИРЗЎўнҳгK»тҒц·ҘМжДбВ“(liЁўn)әПК№УГҢҰCal-62јҡ°ыөДУ°н‘ЎЈ
NЈ©Х№КҫФЪ»щТт№ӨіМРЎКуЦРNSUN2ТЦЦЖ„©Ўў¶аИбұИРЗЎўнҳгKәНҒц·ҘМжДбөДҪoЛҺ·Ҫ°ёКҫТвҲDЎЈ
OЈ©ЙўьcҲDп@КҫёчҪMРЎКуөДЧоҪKД[БцЦШБҝЎЈ
PЈ©ЙўьcҲDп@КҫёчҪMРЎКуөДЧоҪKД[Бцуw·eЎЈ
QЈ©п@КҫёчҪMATCРЎКуөДЙъҙжЗъҫҖЎЈ

У‘Х“әНҫЦПЮРФ
ұҫСРҫҝҪТКҫБЛNSUN2НЁЯ^m5CРЮп—Х{(diЁӨo)№қ(jiЁҰ)SRSF6өДјҡ°ыәЛЩ|(zhЁ¬)ЮD(zhuЁЈn)Я\Ј¬ЯM¶шУ°н‘UAP1јфЗРәНN-МЗ»щ»ҜЛ®ЖҪЈ¬ЧоҪKҢ§(dЁЈo)ЦВATCДНЛҺРФөДҷCЦЖЎЈСРҫҝҪY(jiЁҰ)№ыұнГчЈ¬NSUN2ј°ЖдПВУО°РьcКЗ°РПтATCДНЛҺРФөДқ“ФЪЦОҜҹ°РьcЎЈ
И»¶шЈ¬NSUN2ФЪХэіЈјҡ°ыЙъАнЦРТІ°l(fЁЎ)“]ЦШТӘЧчУГЈ¬ПөҪy(tЁҜng)РФТЦЦЖNSUN2ҝЙДЬ•юҺ§Ғн°ІИ«РФҶ–о}ЎЈҙЛНвЈ¬ИзәОҢҚ¬F(xiЁӨn)NSUN2ТЦЦЖ„©өДД[БцМШ®җРФЯfЛНИФИ»КЗТ»ӮҖМф‘р(zhЁӨn)ЎЈ
ТЧРЎҪY(jiЁҰ)
ұҫСРҫҝНЁЯ^m5C MeRIP-seqәНRNA-seqөИ·ЦОцҪТКҫБЛNSUN2/SRSF6/UAP1РЕМ–ЭSФЪATCДНЛҺРФЦРөДЧчУГҷCЦЖЈ¬һйй_°l(fЁЎ)РВөДЦОҜҹІЯВФМṩБЛАнХ“ТА“ю(jЁҙ)ЎЈNSUN2ТЦЦЖ„©Еc¬F(xiЁӨn)УРҝ№°©ЛҺОпөДВ“(liЁўn)әПК№УГУРНыіЙһйҝЛ·юATCДНЛҺРФөДРВ·Ҫ·ЁЎЈОҙҒнөДСРҫҝРиТӘЯMТ»ІҪФu№АNSUN2ТЦЦЖ„©өД°ІИ«РФәНУРР§РФЈ¬ІўМҪЛчЖдФЪЖдЛыД[БцоҗРНЦРөДқ“ФЪ‘Ә(yЁ©ng)УГЎЈ
m5C MeRIP-seqФЪұҫСРҫҝЦРөДЦШТӘЧчУГ
ұҫСРҫҝөДm5C MeRIP-seqЈЁјЧ»щ»ҜRNAГвТЯіБөнңyРтЈ©ЦчТӘУГУЪҪТКҫNSUN2ФЪјЧ оПЩОҙ·Ц»Ҝ°©ЈЁATCЈ©ЦРөДЧчУГҷCЦЖЈ¬ЦчТӘұн¬F(xiЁӨn)ФЪЈә
ЈЁ1Ј©иb¶Ёm5CРЮп—өД°Р»щТтЈәНЁЯ^MeRIP-seqјјРg(shЁҙ)ДЬүтПөҪy(tЁҜng)өШ·ЦОцNSUN2ФЪATCјҡ°ыЦРҢҰmRNAөДm5CРЮп—ЗйӣrЎЈСРҫҝ°l(fЁЎ)¬F(xiЁӨn)Ј¬NSUN2ДЬүтМШ®җРФөШРЮп—SRSF6 mRNAЙПөДm5CО»ьcЎЈЯ@Т»°l(fЁЎ)¬F(xiЁӨn)һйәуАm(xЁҙ)СРҫҝNSUN2ИзәОНЁЯ^m5CРЮп—Х{(diЁӨo)ҝШ»щТтұнЯ_МṩБЛЦұҪУЧC“ю(jЁҙ)ЎЈ
ЈЁ2Ј©ҪТКҫm5CРЮп—өД№ҰДЬЈәMeRIP-seq·ЦОцп@КҫЈ¬NSUN2ҪйҢ§(dЁЈo)өДm5CРЮп—ДЬүтХ{(diЁӨo)№қ(jiЁҰ)SRSF6 mRNAөДјҡ°ыәЛЩ|(zhЁ¬)ЮD(zhuЁЈn)Я\ЎЈҫЯуwҒнХfЈ¬m5CРЮп—ФцҸҠБЛSRSF6 mRNAҸДјҡ°ыәЛПтјҡ°ыЩ|(zhЁ¬)өДЮD(zhuЁЈn)Я\Ј¬ҸД¶шҙЩЯMБЛSRSF6ө°°ЧөДұнЯ_ЎЈЯ@Т»Я^іМҢҰУЪATCјҡ°ыөД¶аЛҺДНЛҺРФЈЁMDRЈ©ЦБкP(guЁЎn)ЦШТӘЎЈ
ЈЁ3Ј©МҪЛчm5CРЮп—өДХ{(diЁӨo)ҝШҷCЦЖЈәСРҫҝЦРНЁЯ^MeRIP-seqҪY(jiЁҰ)әПЖдЛыҢҚтһКЦ¶ОЈЁИзRNAПВАӯҢҚтһәНRIP-qPCRЈ©Ј¬ҪТКҫБЛALYREFЧчһйm5CРЮп—өД“reader”Ј¬ДЬүтиb¶ЁІўҪY(jiЁҰ)әПm5CРЮп—өДSRSF6 mRNAЈ¬ІўҢўЖдҸДјҡ°ыәЛЮD(zhuЁЈn)Я\өҪјҡ°ыЩ|(zhЁ¬)ЎЈЯ@Т»°l(fЁЎ)¬F(xiЁӨn)ЯMТ»ІҪНкЙЖБЛm5CРЮп—ФЪ»щТтұнЯ_Х{(diЁӨo)ҝШЦРөД·ЦЧУҷCЦЖЎЈ
ЈЁ4Ј©тһЧCm5CРЮп—өДЙъОпҢW(xuЁҰ)ТвБxЈәНЁЯ^ұИЭ^NSUN2ЗГіэјҡ°ыЕcТ°ЙъРНјҡ°ыөДMeRIP-seq”ө(shЁҙ)“ю(jЁҙ)Ј¬СРҫҝИЛҶT°l(fЁЎ)¬F(xiЁӨn)NSUN2ЗГіэәуSRSF6 mRNAөДm5CРЮп—Л®ЖҪп@ЦшПВҪөЈ¬Н¬•rSRSF6ө°°ЧұнЯ_ТІКЬөҪТЦЦЖЎЈЯ@ұнГчm5CРЮп—ФЪҫSіЦSRSF6№ҰДЬәНATCјҡ°ыДНЛҺРФЦР°l(fЁЎ)“]ЦшЦШТӘЧчУГЎЈ
…ўҝјОД«IЈә
Hou X, Dong Q, Hao J, Liu M, Ning J, Tao M, Wang Z, Guo F, Huang D, Shi X, Gao M, Li D, Zheng X. NSUN2-mediated m5C modification drives alternative splicing reprogramming and promotes multidrug resistance in anaplastic thyroid cancer through the NSUN2/SRSF6/UAP1 signaling axis. Theranostics 2025; 15(7):2757-2777. doi:10.7150/thno.104713.
ҪьИХЈ¬МмҪтбt(yЁ©)ҝЖҙуҢW(xuЁҰ)Д[Бцбt(yЁ©)ФәаҚПтЗ°ҪМКЪҲFк ТФјЧ оПЩОҙ·Ц»Ҝ°©ЈЁATCЈ©һйСРҫҝҢҰПуЈ¬АыУГm5C MeRIP-seqөИјјРg(shЁҙ)ҪТКҫБЛNSUN2ҪйҢ§(dЁЈo)өДm5CРЮп—ФЪATCДНЛҺРФЦРөДЧчУГҷCЦЖЈ¬кUГчБЛNSUN2/SRSF6/UAP1РЕМ–ЭSФЪATC¶аЛҺДНЛҺРФЈЁMultidrug Resistance, MDRЈ©ЦРөДкP(guЁЎn)жIЧчУГЈ¬ІўМбіцНЁЯ^РЎ·ЦЧУNSUN2ТЦЦЖ„©ҝЛ·юДНЛҺРФөДРВІЯВФЎЈЯ@Т»°l(fЁЎ)¬F(xiЁӨn)І»ғHһйATCөДЦОҜҹМṩБЛРВөД°РьcЈ¬ТІһйАнҪвRNAРЮп—ФЪД[БцДНЛҺРФЦРөДЧчУГМṩБЛЦШТӘТА“ю(jЁҙ)ЎЈПакP(guЁЎn)СРҫҝіЙ№ыТФЎ¶NSUN2-mediated m5C modification drives alternative splicing reprogramming and promotes multidrug resistance in anaplastic thyroid cancer through the NSUN2/SRSF6/UAP1 signaling axisЎ·һйо}°l(fЁЎ)ұнУЪЎ¶TheranosticsЎ·лsЦҫЎЈ
ҳЛ(biЁЎo)о}ЈәNSUN2-mediated m5C modification drives alternative splicing reprogramming and promotes multidrug resistance in anaplastic thyroid cancer through the NSUN2/SRSF6/UAP1 signaling axisЈЁNSUN2ҪйҢ§(dЁЈo)өДm5CРЮп—НЁЯ^NSUN2/SRSF6/UAP1РЕМ–ЭSтҢ(qЁұ)„УҝЙЧғјфЗРЦШҫҺіМІўҙЩЯMјЧ оПЩОҙ·Ц»Ҝ°©өД¶аЛҺДНЛҺРФЈ©
°l(fЁЎ)ұн•rйgЈә2025-1-27
°l(fЁЎ)ұнЖЪҝҜЈәTheranostics
У°н‘ТтЧУЈәIF 12.4/Q1
јјРg(shЁҙ)ЖҪЕ_ЈәMeRIP-seqЎўscRNA-seqЎўbulk RNA-seqЈЁТЧ»щТтҪрЕЖјјРg(shЁҙ)Ј©
ұҫСРҫҝНЁЯ^ҢҰATCҳУұҫөДbulkЮD(zhuЁЈn)дӣҪMЈЁbulk RNA-seqЈ©әНҶОјҡ°ыRNAңyРтЈЁscRNA-seqЈ©”ө(shЁҙ)“ю(jЁҙ)ЯMРРИ«Гж·ЦОцЈ¬ТФәYЯxЕc¶аЛҺДНЛҺРФЈЁMDRЈ©ПакP(guЁЎn)өДm5CРЮп—»щТтЎЈлSәуЯMРРIC50ҢҚтһЎўБчКҪјҡ°ыРg(shЁҙ)·ЦОцЈ¬ІўАыУГNsun2»щТтЗГіэөДЧФ°l(fЁЎ)РФјЧ оПЩ°©ЈЁspontaneous tumorigenic ATCЈ©РЎКуДЈРНЈ¬ЧCГчБЛNSUN2ФЪATCЦРҙЩЯMMDRөДЧчУГЎЈһйБЛМҪҫҝNSUN2ҪйҢ§(dЁЈo)өДДНЛҺҷCЦЖЈ¬СРҫҝҳӢ(gЁ°u)ҪЁБЛNSUN2»щТтЗГіэөДATCјҡ°ыПөЈ¬ІўЯMРРЮD(zhuЁЈn)дӣҪMҢW(xuЁҰ)Ўўө°°ЧЩ|(zhЁ¬)ҪMҢW(xuЁҰ)әНMeRIP-seq·ЦОцЎЈҙЛНвЯҖНЁЯ^RNA-seqңyРтәНҝЙЧғјфЗР(alternative splicingЈ¬AS)·ЦОцЈ¬СРҫҝNSUN2ЗГіэәуөДХыуwЧғ»ҜЎЈЯMТ»ІҪНЁЯ^МЗө°°ЧИҫЙ«ЎўЧғРФГвТЯіБөн·әЛШ»ҜЎўәЛЩ|(zhЁ¬)·ЦлxәНPCRөИ·Ҫ·ЁЈ¬МҪУ‘БЛNSUN2/SRSF6/UAP1ЭSөДқ“ФЪҷCЦЖЎЈЧоәуФЪуwНвәНуwғИ(nЁЁi)Фu№АБЛРЎ·ЦЧУNSUN2ТЦЦЖ„©Еcҝ№°©ЛҺОпөД…f(xiЁҰ)Н¬Р§‘Ә(yЁ©ng)ЎЈ
СРҫҝҪY(jiЁҰ)№ыұнГчЈ¬NSUN2ұнЯ_ЕcATCЦРөДMDRп@ЦшПакP(guЁЎn)ЎЈNSUN2Ччһйm5CөД“writer”Ј¬ALYREFЧчһй“reader”Ј¬ЛьӮғ№ІН¬ЧчУГУЪSRSF6 mRNAЈ¬ХTҢ§(dЁЈo)ҝЙЧғјфЗРЦШҫҺіМЈ¬ІўҢўUAP1»щТтөДјфҪУРОКҪҸДAGX1ЮD(zhuЁЈn)ПтAGX2ЎЈҪY(jiЁҰ)№ыұнГчЈ¬AGX2ФцҸҠБЛABCЮD(zhuЁЈn)Я\ө°°ЧөДN-МЗ»щ»ҜЈ¬НЁЯ^ТЦЦЖ·әЛШ»ҜҪйҢ§(dЁЈo)өДҪөҪвТФ·Җ(wЁ§n)¶ЁЯ@Р©ө°°ЧЎЈNSUN2ТЦЦЖ„©НЁЯ^ҪөөНNSUN2Гё»оРФІўңpЙЩПВУО°РҳЛ(biЁЎo)ұнЯ_Ј¬һйҝЛ·юATCЦРөДMDRМṩБЛТ»·NРВөДЎўУРЗ°ҫ°өДЦОҜҹІЯВФЎЈ
ұҫСРҫҝҪТКҫБЛNSUN2/SRSF6/UAP1РЕМ–ЭSФЪATCөДMDRЦРөДкP(guЁЎn)жIЧчУГЈ¬Іўҙ_¶ЁБЛNSUN2Ччһй»ҜҜҹәН°РПтЦОҜҹөД…f(xiЁҰ)Н¬°РьcЎЈ
СРҫҝХӘТӘ
СРҫҝ·Ҫ·Ё
ЙъОпРЕПўҢW(xuЁҰ)·ЦОцЈәНЁЯ^·ЦОцATCҳУұҫөДbulk RNA-seqңyРт”ө(shЁҙ)“ю(jЁҙ)Ј¬әYЯxЕcMDRПакP(guЁЎn)өДm5CРЮп—»щТтЎЈ
јҡ°ыҢҚтһәН·ЦЧУҷCЦЖСРҫҝЈәАыУГCRISPR/Cas9јјРg(shЁҙ)ҳӢ(gЁ°u)ҪЁNSUN2»щТтЗГіэЈЁknockoutЈ©өДATCјҡ°ыПөЈ¬НЁЯ^RNA-seqЎўө°°ЧЩ|(zhЁ¬)ҪMҢW(xuЁҰ)әНMeRIP-seq·ЦОцөИјјРg(shЁҙ)СРҫҝNSUN2өД№ҰДЬЎЈ
„УОпДЈРНЈәҳӢ(gЁ°u)ҪЁNsun2»щТтЗГіэөДЧФ°l(fЁЎ)РФјЧ оПЩ°©РЎКуДЈРНЈ¬Фu№АNSUN2уwғИ(nЁЁi)ҢҰMDRУ°н‘ЎЈ
ЛҺОпГфёРРФ·ЦОцЈәНЁЯ^IC50ҢҚтһәНБчКҪјҡ°ыРg(shЁҙ)ҷzңyNSUN2ТЦЦЖ„©Еcҝ№°©ЛҺОпөДВ“(liЁўn)әПР§‘Ә(yЁ©ng)ЎЈ
СРҫҝҪY(jiЁҰ)№ы
ЈЁ1Ј©ЙъОпРЕПўҢW(xuЁҰ)·ЦОцҪТКҫATCЦРNSUN2әН¶аЛҺДНЛҺРФЈЁMDRЈ©Ц®йgөДПакP(guЁЎn)РФ
НЁЯ^·ЦОцCancer Therapeutics Response PortalЈЁCTRPЈ©”ө(shЁҙ)“ю(jЁҙ)ҺмЦРөДЛҺОпГфёРРФ”ө(shЁҙ)“ю(jЁҙ)Ј¬°l(fЁЎ)¬F(xiЁӨn)NSUN2өДұнЯ_Еc¶а·Nҝ№°©ЛҺОпЈЁ°ьАЁ»ҜҜҹЛҺОпәНАТ°ұЛбјӨГёТЦЦЖ„©Ј©өДIC50ЦөіКХэПакP(guЁЎn)ЎЈ
АыУГGEO”ө(shЁҙ)“ю(jЁҙ)ҺмЦРөДATCЮD(zhuЁЈn)дӣҪM”ө(shЁҙ)“ю(jЁҙ)Ј¬Ңў»јХЯ·ЦһйёЯNSUN2ұнЯ_ҪMәНөНNSUN2ұнЯ_ҪMЈ¬°l(fЁЎ)¬F(xiЁӨn)ёЯNSUN2ұнЯ_Еcјҡ°ы·ЦБСЎўУРҪz·ЦБСЎўјҡ°ыЦЬЖЪөИ»ҜҜҹ°РПтЯ^іМПакP(guЁЎn)ЎЈ
scRNA-seqңyРт·ЦОцп@КҫЈ¬NSUN2ФЪATCјҡ°ыЦРөДұнЯ_Еc¶а·N»ҜҜҹЛҺОпәНTKIөДДНЛҺРФПакP(guЁЎn)ЎЈ
ҲD1ЈәЙъОпРЕПўҢW(xuЁҰ)·ЦОцҪТКҫNSUN2ЕcATCЦР¶аЛҺДНЛҺРФЈЁMDRЈ©өДПакP(guЁЎn)РФЎЈ
B-CЈ©ёЯNSUN2ұнЯ_ҪMЕcөНNSUN2ұнЯ_ҪMЦ®йgІо®җұнЯ_»щТтЈЁDEGsЈ©өДё»јҜНЁВ·ҝӮҪY(jiЁҰ)ЎЈ
DЈ©ATCөДscRNA-seq”ө(shЁҙ)“ю(jЁҙ)өДtSNEҲDЈ¬Х№КҫБЛиb¶ЁіцөДјҡ°ыclustersЈЁn=10Ј©ЎЈ
EЈ©ЙўьcҲDп@КҫNSUN2к–РФјҡ°ыЕcNSUN2кҺРФјҡ°ыөДlog2ұ¶Чғ»ҜЈЁlog2FCЈ©ј°ЖдЕcГҝ·NЛҺОпоA(yЁҙ)ңyөДIC50ЗъҫҖПВГж·eЈЁAUCЈ©ЦөөДПакP(guЁЎn)РФЎЈ
FЈ©әыөыҲDХ№КҫNSUN2ұнЯ_ЎўЛҺОпГфёРРФЕc»щТтұҫуwЈЁGOЈ©НЁВ··ЦОцЦ®йgөДПакP(guЁЎn)РФЎЈ
GЈ©ATCөДscRNA-seq”ө(shЁҙ)“ю(jЁҙ)өДtSNEҲDЈ¬Х№КҫБЛиb¶ЁіцөДјҡ°ыclustersЈЁn=10Ј©ЎЈ
HЈ©К№УГBeyondcellҢҰGEO”ө(shЁҙ)“ю(jЁҙ)јҜҳУұҫЯMРРөДҶОјҡ°ыЛҺОпГфёРРФ·ЦОцЎЈ
IЈ©NSUN2ұнЯ_ЕcBeyondcellФu·ЦЦ®йgөДПакP(guЁЎn)РФЎЈ
ЈЁ2Ј©ATCЦРNSUN2ХTҢ§(dЁЈo)өДMDRИЎӣQУЪЖдјЧ»щЮD(zhuЁЈn)ТЖГё»оРФ
ФЪATCјҡ°ыПөЦРЗГөН»тЗГіэNSUN2әуЈ¬јҡ°ыҢҰ¶а·NЛҺОпөДГфёРРФп@ЦшФцјУЈ¬ұнГчNSUN2ұнЯ_ЕcMDRГЬЗРПакP(guЁЎn)ЎЈ
НЁЯ^ҳӢ(gЁ°u)ҪЁNSUN2ҙЯ»Ҝ»оРФН»ЧғуwЈЁC271AЈ©Ј¬°l(fЁЎ)¬F(xiЁӨn)Ц»УРТ°ЙъРНNSUN2ДЬүт»ЦҸН(fЁҙ)m5CРЮп—»оРФәНЛҺОпДНКЬРФЈ¬ұнГчNSUN2өДјЧ»щЮD(zhuЁЈn)ТЖГё»оРФКЗЖдХTҢ§(dЁЈo)MDRөДкP(guЁЎn)жIЎЈ
ҲD2ЈәATCЦРУЙNSUN2ХTҢ§(dЁЈo)өДMDRТАЩҮУЪЖдјЧ»щЮD(zhuЁЈn)ТЖГё»оРФЎЈ
AЈ©НЁЯ^Western blot·ЦОцҷzңyCal-62јҡ°ыЦРNSUN2»щТтЗГіэәуөДNSUN2ө°°ЧұнЯ_ЗйӣrЎЈBЈ©НЁЯ^ьcлsҪ»ҢҚтһҷzңyNSUN2»щТтЗГіэҢҰCal-62јҡ°ыmRNAЮD(zhuЁЈn)дӣҪMЦРm5CЛ®ЖҪөДУ°н‘ЎЈ
CЈ©НЁЯ^ұИЙ«·Ёm5C¶ЁБҝҢҚтһҙ_ХJNSUN2»щТтЗГіэәуCal-62јҡ°ыЦРm5CЛ®ЖҪөДЧғ»ҜЎЈ
D-FЈ©НЁЯ^јҡ°ыЙъйLТЦЦЖҢҚтһФu№АNSUN2»щТтЗГіэҢҰCal-62јҡ°ыҢҰ¶аИбұИРЗЈЁdoxorubicinЈ©ЎўнҳгKЈЁcisplatinЈ©әНҒц·ҘМжДбЈЁlenvatinibЈ©ГфёРРФөДУ°н‘ЎЈ
GЈ©НЁЯ^Western blot·ЦОцҷzңyCal-62јҡ°ыҪӣ(jЁ©ng)І»Н¬МҺАнәуNSUN2ө°°ЧөДПаҢҰұнЯ_БҝЎЈ
HЈ©НЁЯ^ьcлsҪ»ҢҚтһҷzңyҪӣ(jЁ©ng)ЭdуwМҺАнәуCal-62јҡ°ыmRNAЮD(zhuЁЈn)дӣҪMЦРm5CШS¶ИЎЈ
IЈ©НЁЯ^ұИЙ«·Ёm5C¶ЁБҝҢҚтһҙ_ХJҪӣ(jЁ©ng)МҺАнәуCal-62јҡ°ыЦРm5CЛ®ЖҪЧғ»ҜЎЈ
J-LЈ©НЁЯ^јҡ°ыЙъйLТЦЦЖҢҚтһФu№АҪӣ(jЁ©ng)І»Н¬ЭdуwЮD(zhuЁЈn)ИҫөДCal-62јҡ°ыөДIC50ЦөЎЈ
MЈ©ЧФ°l(fЁЎ)РФATCРЎКуДЈРНөДҳӢ(gЁ°u)ҪЁ·Ҫ°ёЈ¬°ьАЁ°РПтІЯВФЈЁЧуЈ©әН·ұЦі·Ҫ°ёЈЁУТЈ©ЎЈ
NЈ©РЎКу»щТт·ЦРНҪY(jiЁҰ)№ыЈ¬ЖдЦРTPO-creөДDNAДэДz—lҺ§ҙуРЎһй130/324bpЈ¬BrafV600Eһй185/307bpЈ¬Trp53flox/floxһй290/370bpЈ¬Nsun2flox/floxһй151/212bpЎЈ
OЈ©ФЪ»щТт№ӨіМРЎКуЦР¶аИбұИРЗЎўнҳгKәНҒц·ҘМжДбөДҪoЛҺ·Ҫ°ёЎЈmATCЈЁРЎКујЧ оПЩОҙ·Ц»Ҝ°©Ј©ЎЈ
PЈ©ЙўьcҲDп@КҫёчҪMРЎКуөДЧоҪKД[БцЦШБҝЎЈ
QЈ©ЙўьcҲDп@КҫёчҪMРЎКуөДЧоҪKД[Бцуw·eЎЈ
RЈ©п@КҫёчҪMATCРЎКуөДЙъҙжЗъҫҖЎЈ
ЈЁ3Ј©ФЪATCЦРЈ¬NSUN2ҪйҢ§(dЁЈo)өДm5CРЮп—НЁЯ^°РПтSRSF6тҢ(qЁұ)„УҝЙЧғјфЗРЦШҫҺіМ
ө°°ЧЩ|(zhЁ¬)ҪMҢW(xuЁҰ)·ЦОцп@КҫЈ¬NSUN2ЗГіэәуЈ¬ЕcRNAјфҪУПакP(guЁЎn)өДө°°ЧұнЯ_°l(fЁЎ)Йъп@ЦшЧғ»ҜЈ¬ЖдЦРSRSF6ө°°ЧұнЯ_п@ЦшПВХ{(diЁӨo)Ј¬¶шЖдmRNAЛ®ЖҪІ»ЧғЎЈ
MeRIP-seqәНqPCRҪY(jiЁҰ)№ыұнГчЈ¬NSUN2НЁЯ^m5CРЮп—Х{(diЁӨo)№қ(jiЁҰ)SRSF6 mRNAөДәЛЩ|(zhЁ¬)ЮD(zhuЁЈn)Я\Ј¬ЯM¶шУ°н‘Ждө°°ЧұнЯ_ЎЈ
НЁЯ^Н»ЧғSRSF6 mRNAЙПөДm5CРЮп—О»ьcЈ¬°l(fЁЎ)¬F(xiЁӨn)Я@Р©О»ьcҢҰSRSF6өДәЛЩ|(zhЁ¬)ЮD(zhuЁЈn)Я\ЦБкP(guЁЎn)ЦШТӘЎЈ
ҲD3ЈәATCЦРЈ¬NSUN2ҪйҢ§(dЁЈo)өДm5CРЮп—НЁЯ^°РПтSRSF6тҢ(qЁұ)„УҝЙЧғјфЗРЦШҫҺіМЎЈ
AЈ©НЁЯ^TMT-MSҷzңyөДCal-62јҡ°ыЦРNSUN2»щТтЗГіэәуө°°ЧЛ®ЖҪЧғ»ҜөД»рЙҪҲDЎЈBЈ©Cal-62јҡ°ыЦРNSUN2»щТтЗГіэәуTMT-MS”ө(shЁҙ)“ю(jЁҙ)өДGOё»јҜ·ЦОцЎЈ
CЈ©Cal-62јҡ°ыЦРNSUN2»щТтЗГіэәуІо®җјфҪУ»щТтөД°Щ·ЦұИјфҪУ°ьә¬ЈЁΔPSIЈ©өД»рЙҪҲDЎЈ
DЈ©Cal-62јҡ°ыЦРNSUN2»щТтЗГіэәуп@ЦшΔPSIЧғ»ҜөДРЎМбЗЩҲDЎЈ
EЈ©Cal-62јҡ°ыЦРNSUN2»щТтЗГіэәуSF3A/BЎўU2AFәЛРДҸН(fЁҙ)әПуwәНhnRNPјТЧеЦРDEGҹбҲDЎЈ
FЈ©Cal-62јҡ°ыЦРNSUN2»щТтЗГіэәуTMT-MS”ө(shЁҙ)“ю(jЁҙ)ЦРSRSF6ө°°ЧЛ®ЖҪөДҪy(tЁҜng)УӢ·ЦОцЎЈ
GЈ©Cal-62јҡ°ыЦРNSUN2»щТтЗГіэәуЦёКҫө°°ЧөДWestern blot·ЦОцЎЈ
HЈ©Cal-62јҡ°ыЦРNSUN2»щТтЗГіэәуSRSF6»щТтЧщЦЬҮъөДMeRIP-seq”ө(shЁҙ)“ю(jЁҙ)өДIGVЬүЫEҲDЎЈ
IЈ©Cal-62јҡ°ыЦРNSUN2»щТтЗГіэәуSRSF6 mRNAЦРm5Cё»јҜөДMeRIP-qPCRҷzңyЎЈ
JЈ©Ҫӣ(jЁ©ng)ЦёКҫЭdуwМҺАнөДCal-62јҡ°ыЦРSRSF6 mRNAЦРm5Cё»јҜөДMeRIP-qPCR·ЦОцЎЈ
KЈ©NSUN2»щТтЗГіэәуCal-62јҡ°ыЦРSRSF6 mRNAөДәЛЩ|(zhЁ¬)·ЦІјqRT-PCR·ЦОцЎЈ
LЈ©Ҫӣ(jЁ©ng)ЦёКҫЭdуwМҺАнөДCal-62јҡ°ыЦРSRSF6 mRNAөДәЛЩ|(zhЁ¬)·ЦІјЎЈ
MЈ©Ҫӣ(jЁ©ng)ЦёКҫЭdуwМҺАнөДCal-62јҡ°ыЦРЦёКҫө°°ЧөДWestern blot·ЦОцЎЈ
NЈ©SRSF6ЦРөДөдРНm5CҪY(jiЁҰ)әПО»ьcЎЈн”Іҝһйm5CҪY(jiЁҰ)әПО»ьcөДmotifРтБРЈ»өЧІҝһйSRSF6ЦРиb¶ЁөДm5CО»ьcЎЈ
OЈ©Ҫӣ(jЁ©ng)ЦёКҫЭdуwМҺАнөДCal-62јҡ°ыЦРSRSF6 mRNAЦРm5Cё»јҜөДMeRIP-qPCR¶ЁБҝЎЈ
PЈ©Ҫӣ(jЁ©ng)ЦёКҫЭdуwМҺАнөДCal-62јҡ°ыЦРSRSF6 mRNAөДәЛЩ|(zhЁ¬)·ЦІјЎЈ
ЈЁ4Ј©NSUN2НЁЯ^ФцҸҠABCЮD(zhuЁЈn)Я\ө°°ЧөДN-МЗ»щ»ҜҙЩЯMATCЦРөДMDR
ЮD(zhuЁЈn)дӣҪM·ЦОцәНKEGGНЁВ··ЦОцп@КҫЈ¬NSUN2…ўЕcN-МЗжңЙъОпәПіЙЯ^іМЎЈ
НЁЯ^МЗө°°ЧИҫЙ«әНө°°ЧЩ|(zhЁ¬)УЎЫE·ЦОцЈ¬°l(fЁЎ)¬F(xiЁӨn)NSUN2ЗГіэәуЈ¬ABCЮD(zhuЁЈn)Я\ө°°ЧЈЁABCC1әНABCG2Ј©өДN-МЗ»щ»ҜЛ®ЖҪҪөөНЈ¬ө°°Ч·Җ(wЁ§n)¶ЁРФПВҪөЎЈ
БчКҪјҡ°ыРg(shЁҙ)ҷzңyп@КҫЈ¬NSUN2ЗГіэјҡ°ыҢҰЛҺОп”zИЎФцјУЈ¬ұнГчЛҺОпНвЕЕңpЙЩЈ¬ДНЛҺРФҪөөНЎЈ
ҲD4ЈәNSUN2НЁЯ^ФцҸҠABCЮD(zhuЁЈn)Я\ө°°ЧөДN-МЗ»щ»ҜҙЩЯMATCЦРөДMDRЎЈ
AЈ©Cal-62јҡ°ыЦРNSUN2»щТтЗГіэәуЮD(zhuЁЈn)дӣ”ө(shЁҙ)“ю(jЁҙ)өДGOё»јҜ·ЦОцЎЈB-CЈ©Cal-62јҡ°ыЦРNSUN2»щТтЗГіэәуЮD(zhuЁЈn)дӣ”ө(shЁҙ)“ю(jЁҙ)өД»щТтјҜё»јҜ·ЦОцЈЁGSEAЈ©ЎЈ
DЈ©ҝӮө°°ЧөДҝјсRЛ№ББЛ{ИҫЙ«ЈЁЧуЈ©Ј¬NSUN2»щТтЗГіэәуCal-62јҡ°ыөДМЗө°°ЧИҫЙ«ҪY(jiЁҰ)№ыЈЁУТЈ©ЎЈ
E-FЈ©Ҫӣ(jЁ©ng)»тОҙҪӣ(jЁ©ng)TMЈЁtunicamycinЈ©ЎўSSЈЁswainsonineЈ©МҺАнөДCal-62јҡ°ыЦРЦёКҫө°°ЧЛ®ЖҪөДWestern blot·ЦОцЎЈјҡ°ыБСҪвҳУұҫҪӣ(jЁ©ng)»тОҙҪӣ(jЁ©ng)PNGase FМҺАнЎЈ“G”ұнКҫN-МЗ»щ»ҜЈ¬“DG”ұнКҫN-МЗ»щ»ҜңpЙЩЎЈ
GЈ©NSUN2»щТтЗГіэөДCal-62јҡ°ыЦРЦёКҫө°°ЧЛ®ЖҪөДWestern blot·ЦОцЎЈ
HЈ©ҙъұнРФГвТЯҪM»ҜЈЁIHCЈ©ҲDПсп@Кҫ35АэATCҪMҝ—ЦРNSUN2ЎўABCG2әНABCC1өДұнЯ_ЗйӣrЎЈ
I-JЈ©ІЙУГҝЁ·Ҫҷzтһ·ЦОцNSUN2ЕcABCG2»тABCC1ұнЯ_Ц®йgөДПакP(guЁЎn)РФЎЈ
K-LЈ©NSUN2ЕcABCG2»тABCC1өДIHCФu·ЦөДПакP(guЁЎn)РФ·ЦОцЎЈ
M-NЈ©К№УГNetNglyc·ю„Х(wЁҙ)ЖчоA(yЁҙ)ңyөДABCG2әНABCC1өДқ“ФЪN-МЗ»щ»ҜМ춬хЈ°·О»ьcЎЈ
O-PЈ©·Ц„eФЪЮD(zhuЁЈn)ИҫБЛN557QЎўN388QЎўN596Q»тN71QЎўN19QЎўN310QЎўN(71+310)QН»ЧғЩ|(zhЁ¬)БЈөДCal-62јҡ°ыЦРЈ¬НЁЯ^Western blotҷzңyABCG2әНABCC1өДұнЯ_ЗйӣrЎЈ
Q-SЈ©NSUN2»щТтЗГіэәуCal-62јҡ°ыЦР¶аИбұИРЗЎўнҳгKәНҒц·ҘМжДбөДјҡ°ығИ(nЁЁi)·eАЫЗйӣrөДБчКҪјҡ°ығx·ЦОцЎЈУТӮИ(cЁЁ)п@КҫЖҪҫщҹЙ№вҸҠ¶ИөД¶ЁБҝҪY(jiЁҰ)№ыЎЈ
ЈЁ5Ј©NSUN2НЁЯ^N-МЗ»щ»ҜФцҸҠABCЮD(zhuЁЈn)Я\ө°°ЧөД·Җ(wЁ§n)¶ЁРФ
НЁЯ^western blot ·ЦОцәН·әЛШ»ҜҢҚтһЈ¬°l(fЁЎ)¬F(xiЁӨn)NSUN2ЗГіэәуЈ¬ABCC1әНABCG2өД·әЛШ»ҜЛ®ЖҪФцјУЈ¬ұнГчN-МЗ»щ»ҜИұК§Ң§(dЁЈo)ЦВЯ@Р©ө°°ЧёьИЭТЧұ»ҪөҪвЎЈ
К№УГө°°ЧГёуwТЦЦЖ„©MG132ҝЙТФІҝ·Ц»ЦҸН(fЁҙ)ABCC1әНABCG2өДө°°ЧЛ®ЖҪЈ¬ЯMТ»ІҪЧCҢҚБЛN-МЗ»щ»ҜҢҰө°°Ч·Җ(wЁ§n)¶ЁРФөДУ°н‘ЎЈ
ҲD5ЈәNSUN2НЁЯ^N-МЗ»щ»ҜФцҸҠABCЮD(zhuЁЈn)Я\ө°°ЧөД·Җ(wЁ§n)¶ЁРФЎЈ
AЈ©Cal-62јҡ°ыҪӣ(jЁ©ng)TM»тІ»Н¬қв¶ИөДMG132МҺАнәуЈ¬К№УГЦё¶Ёҝ№уwЯMРРWestern blot·ЦОцЎЈBЈ©NSUN2»щТтЗГіэөДCal-62јҡ°ыҪӣ(jЁ©ng)І»Н¬қв¶ИөДMG132МҺАнәуЈ¬К№УГЦё¶Ёҝ№уwЯMРРWestern blot·ЦОцЎЈ
CЈ©Ҫӣ(jЁ©ng)TMәНMG132МҺАнөДCal-62јҡ°ыЦРЦёКҫө°°ЧЛ®ЖҪөДWestern blot·ЦОцЎЈ
DЈ©К№УГImageJЬӣјюҢҰABCC1әНABCG2ө°°ЧЛ®ЖҪЯMРР¶ЁБҝ·ЦОцЎЈ
E-FЈ©Ҫӣ(jЁ©ng)TMәНMG132МҺАнөДCal-62јҡ°ыЯMРРABCC1»тABCG2ГвТЯіБөнЈЁIPЈ©Ј¬лSәуК№УГҝ№·әЛШҝ№уwЯMРРWestern blot·ЦОцЎЈ
GЈ©NSUN2»щТтЗГіэЗТҪӣ(jЁ©ng)MG132МҺАнөДCal-62јҡ°ыЦРЦёКҫө°°ЧЛ®ЖҪөДWestern blot·ЦОцЎЈ
HЈ©К№УГImageJЬӣјюҢҰABCC1әНABCG2ө°°ЧҸҠ¶ИЯMРР¶ЁБҝ·ЦОцЎЈ
I-JЈ©NSUN2»щТтЗГіэЗТҪӣ(jЁ©ng)MG132МҺАнөДCal-62јҡ°ыЯMРРABCC1»тABCG2ГвТЯіБөнЈЁIPЈ©Ј¬лSәуК№УГҝ№·әЛШҝ№уwЯMРРWestern blot·ЦОцЎЈ
K-LЈ©ФЪCal-62јҡ°ыЦРұнЯ_өДТ°ЙъРНЈЁWTЈ©»тЦёКҫөДABCG2/ABCC1Н»ЧғуwҪӣ(jЁ©ng)125μg/mLӯh(huЁўn)јәхЈҒҶ°·ЈЁCHXЈ©МҺАнМШ¶Ё•rйgйgёфәуЈ¬НЁЯ^Western blotЯMРР·ЦОцЎЈ
M-NЈ©К№УГImageJЬӣјюҢҰABCG2»тABCC1өДө°°ЧҸҠ¶И¶ЁБҝ·ЦОцЈ¬ІўТФβ-ActinЛ®ЖҪҡwТ»»ҜЎЈ
ЈЁ6Ј©NSUN2НЁЯ^ҪйҢ§(dЁЈo)UAP1өҪSRSF6өДҝЙЧғјфЗРҒнҙЩЯMN-МЗ»щ»Ҝ
UAP1КЗN-МЗ»щ»Ҝ·ҙ‘Ә(yЁ©ng)ЦРUDP-GlcNAcәПіЙөДкP(guЁЎn)жIГёЈ¬ЖдУРғЙ·NҒҶРНЈЁAGX1әНAGX2Ј©ЎЈ
NSUN2ЗГіэәуЈ¬UAP1өДјфҪУРОКҪҸДAGX2ЮD(zhuЁЈn)Чғ?yЁӯu)йAGX1Ј¬Ң§(dЁЈo)ЦВN-МЗ»щ»Ҝ»оРФҪөөНЎЈ
НЁЯ^Я^ұнЯ_SRSF6Ј¬ҝЙТФДжЮD(zhuЁЈn)Я@Т»јфҪУЧғ»ҜЈ¬»ЦҸН(fЁҙ)N-МЗ»щ»ҜЛ®ЖҪәНЛҺОпДНКЬРФЎЈ
ҲD6ЈәNSUN2НЁЯ^SRSF6ҪйҢ§(dЁЈo)UAP1өДҝЙЧғјфЗРҙЩЯMN-МЗ»щ»ҜЎЈ
AЈ©UAP1ҙЩЯMUDP-GlcNAcөДЙъОпәПіЙЈ¬UDP-GlcNAcКЗМЗ»щ»Ҝ·ҙ‘Ә(yЁ©ng)ЦРөДкP(guЁЎn)жI№©уwЈ¬УИЖдФЪN-МЗ»щ»ҜәНO-GlcNAcРЮп—ЦРЎЈBЈ©SashimiҲDп@КҫNSUN2»щТтЗГіэәуCal-62јҡ°ыЦРUAP1јфЗРКВјюөДЧғ»ҜЎЈ
CЈ©ҢҰХХҪMЕcNSUN2»щТтЗГіэөДCal-62јҡ°ыЦРUAP1јфЗРөДRT-PCR·ЦОцЎЈ
DЈ©ЧФ°l(fЁЎ)РФРЎКуATCД[БцЦРUAP1өДјфЗРКВјюөДRT-PCRЈ¬УР»тӣ]УРNsun2»щТтЗГіэЎЈ
EЈ©Ҫӣ(jЁ©ng)ЦёКҫЭdуwМҺАнөДCal-62јҡ°ыЦРUAP1јфЗРКВјюөДRT-PCRЈЁЙПҲDЈ©ЎЈWestern blot·ЦОцБЛSRSF6ЦШҪЁЈЁПВҲDЈ©ЎЈ
FЈ©ҷzңyҝӮө°°ЧөДҝјсRЛ№ББЛ{ИҫЙ«ЈЁЧуЈ©ЎЈҪӣ(jЁ©ng)ЦёКҫЭdуwМҺАнөДCal-62јҡ°ыМЗө°°ЧИҫЙ«ЈЁУТЈ©ЎЈ
GЈ©Ҫӣ(jЁ©ng)ЦёКҫЭdуwМҺАнөДCal-62јҡ°ыЦРЦёКҫө°°ЧЛ®ЖҪөДWestern blot·ЦОцЎЈ
H-JЈ©Ҫӣ(jЁ©ng)ЦёКҫЭdуwМҺАнөДCal-62јҡ°ыЦР¶аИбұИРЗЎўнҳгKәНҒц·ҘМжДбөДјҡ°ығИ(nЁЁi)·eАЫЗйӣrөДБчКҪјҡ°ығx·ЦОцЎЈУТӮИ(cЁЁ)п@КҫБЛЖҪҫщҹЙ№вҸҠ¶ИөД¶ЁБҝҪY(jiЁҰ)№ыЎЈ
K-MЈ©ЦёКҫЭdуwМҺАнCal-62јҡ°ыҢҰ¶аИбұИРЗЎўнҳгKәНҒц·ҘМжДбГфёРУ°н‘јҡ°ыЙъйLТЦЦЖҢҚтһЎЈ
N-PЈ©ЦёКҫЭdуwМҺАнCal-62јҡ°ыҢҰ¶аИбұИРЗЎўнҳгKәНҒц·ҘМжДб·ҙ‘Ә(yЁ©ng)У°н‘өДД[БцЙъйLЗъҫҖЎЈ
Q-SЈ©ЦёКҫЭdуwМҺАнCal-62јҡ°ыҢҰ¶аИбұИРЗЎўнҳгKәНҒц·ҘМжДб·ҙ‘Ә(yЁ©ng)У°н‘Д[БцЧоҪKЦШБҝЎЈ
ЈЁ7Ј©Ччһйm5CөД“reader”Ј¬ ALYREFҝЙТФиb¶ЁSRSF6ІўҢўЖдҸДјҡ°ыәЛЮD(zhuЁЈn)Я\ЦБјҡ°ыЩ|(zhЁ¬)
НЁЯ^RNAГвТЯіБөнЈЁRIPЈ©әНRNAПВАӯҢҚтһЈ¬°l(fЁЎ)¬F(xiЁӨn)ALYREFДЬүтЦұҪУҪY(jiЁҰ)әПm5CРЮп—өДSRSF6 mRNAЈ¬ІўҢўЖдҸДјҡ°ыәЛЮD(zhuЁЈn)Я\ЦБјҡ°ыЩ|(zhЁ¬)ЎЈ
ALYREFөДK171AН»Чғуwп@ЦшҪөөНҢҰm5C -SRSF6ҪY(jiЁҰ)әПДЬБҰЈ¬ұнГчALYREFКЗm5CРЮп—өД“reader”ЎЈ
ALYREFЗГөН»тЯ^ұнЯ_ҝЙТФХ{(diЁӨo)ҝШSRSF6өДјҡ°ыәЛЩ|(zhЁ¬)ЮD(zhuЁЈn)Я\әНө°°ЧұнЯ_Ј¬ЯM¶шУ°н‘ЛҺОпДНКЬРФЎЈ
ҲD7ЈәALYREFЧчһйm5C “reader”Ј¬ДЬүтиb¶ЁІўҪйҢ§(dЁЈo)SRSF6ҸДјҡ°ыәЛөҪјҡ°ыЩ|(zhЁ¬)өДЮD(zhuЁЈn)Я\ЎЈ
A-BЈ©GSEA·ЦОцұнГчALYREFөДұнЯ_Еc»ҜҜҹДНЛҺРФПакP(guЁЎn)ЎЈCЈ©ҹбҲDп@КҫФЪNSUN2»щТтЗГіэ»тALYREFЎўYBX1әНSRSF2»щТтЗГөНөДCal-62јҡ°ыЦРЕc¶аИбұИРЗәНнҳгKДНЛҺРФПакP(guЁЎn)өД»щТтЎЈ
DЈ©НЁЯ^RIP-qPCRҷzңyALYREFЕcCal-62јҡ°ыЦРSRSF6өДҪY(jiЁҰ)әПЎЈ
EЈ©НЁЯ^Western blot·ЦОцҸДSRSF6ЈЁm5CЈ©ПВАӯҢҚтһЦР«@өГөДЦёКҫө°°ЧЎЈ
FЈ©НЁЯ^RIP-qPCRұИЭ^ФЪҪӣ(jЁ©ng)ЦёКҫЭdуwМҺАнөДCal-62јҡ°ыЦРЈ¬Т°ЙъРНALYREFЈЁALYREF-WTЈ©әНK171AН»ЧғРНALYREFЈЁALYREF-K171AЈ©ЕcSRSF6өДҪY(jiЁҰ)әПЎЈ
GЈ©НЁЯ^RIP-qPCRФu№АNSUN2»щТтЗГіэәуCal-62јҡ°ыЦРALYREFЕcSRSF6 mRNAөДҪY(jiЁҰ)әПЎЈ
HЈ©НЁЯ^RIP-qPCRФu№АҪӣ(jЁ©ng)ЦёКҫЭdуwМҺАнөДCal-62јҡ°ыЦРALYREFЕcSRSF6 mRNAөДҪY(jiЁҰ)әПЎЈ
IЈ©НЁЯ^qRT-PCRҢҚтһФu№АALYREF»щТтЗГөНөДCal-62јҡ°ыЦРSRSF6 mRNAөДәЛЩ|(zhЁ¬)·ЦІјЎЈ
JЈ©ФЪЯ^ұнЯ_ALYREF»тALYREF-K171AН»ЧғуwөДCal-62јҡ°ыЦРЈ¬SRSF6 mRNAөДәЛЩ|(zhЁ¬)·ЦІјЎЈ
KЈ©Ҫӣ(jЁ©ng)ЦёКҫЭdуwМҺАнөДCal-62јҡ°ыЦРSRSF6 mRNAөДәЛЩ|(zhЁ¬)·ЦІјЎЈ
L-NЈ©јҡ°ыЙъйLТЦЦЖҢҚтһФu№АЦёКҫЭdуwМҺАнҢҰCal-62јҡ°ыҢҰ¶аИбұИРЗЎўнҳгKәНҒц·ҘМжДб·ҙ‘Ә(yЁ©ng)өДУ°н‘ЎЈ
OЈ©НЁЯ^Western blot·ЦОцҪӣ(jЁ©ng)ЦёКҫЭdуwМҺАнөДCal-62јҡ°ыЦРЦёКҫө°°ЧөДЛ®ЖҪЎЈ
PЈ©NSUN2»щТтЗГіэөДCal-62јҡ°ыЦРALYREFөДГвТЯҹЙ№вИҫЙ«ЈЁЧуӮИ(cЁЁ)Ј©ТФј°әЛ/Щ|(zhЁ¬)ALYREF·ЦІјөД¶ЁБҝ·ЦОцЈЁУТӮИ(cЁЁ)Ј©ЎЈ
QЈ©НЁЯ^Western blot·ЦОцЈЁЧуӮИ(cЁЁ)Ј©ТФј°Па‘Ә(yЁ©ng)өД¶ЁБҝ·ЦОцЈЁУТӮИ(cЁЁ)Ј©Ј¬Х№КҫҢҰХХҪMәНNSUN2»щТтЗГіэөДCal-62јҡ°ыЦРәЛәНЩ|(zhЁ¬)ALYREFөДЛ®ЖҪЈ¬PARP1әНTUBULIN·Ц„eЧчһйәЛәНЩ|(zhЁ¬)ҳЛ(biЁЎo)УӣЎЈ
ЈЁ8Ј©РЎ·ЦЧУNSUN2ТЦЦЖ„©ФцҸҠATCЦР¶аЛҺГфёРРФ
ФO(shЁЁ)УӢІўәПіЙРЎ·ЦЧУNSUN2ТЦЦЖ„©ЈЁNSUN2iЈ©Ј¬ДЬүтп@ЦшҪөөНATCјҡ°ыЦРөДm5CЛ®ЖҪЎЈ
NSUN2iМҺАнәуЈ¬SRSF6өДәЛЩ|(zhЁ¬)ЮD(zhuЁЈn)Я\КЬЧиЈ¬UAP1јфҪУРОКҪёДЧғЈ¬N-МЗ»щ»ҜЛ®ЖҪҪөөНЈ¬ABCЮD(zhuЁЈn)Я\ө°°Ч·Җ(wЁ§n)¶ЁРФПВҪөЎЈ
уwНвҢҚтһәН„УОпДЈРНҢҚтһҫщп@КҫЈ¬NSUN2iЕc»ҜҜҹЛҺОп»тTKIВ“(liЁўn)әПК№УГҝЙТФп@ЦшФцҸҠЛҺОпөДҝ№Д[БцР§№ыЎЈ
ҲD8ЈәРЎ·ЦЧУNSUN2ТЦЦЖ„©ФцҸҠATCҢҰ¶аЛҺөДГфёРРФЎЈ
AЈ©НЁЯ^ьcлsҪ»ҢҚтһФu№АNSUN2»щТтЗГіэөДҪӣ(jЁ©ng)»тОҙҪӣ(jЁ©ng)NSUN2ТЦЦЖ„©МҺАнөДCal-62јҡ°ыЦРmRNAөДm5CЛ®ЖҪЎЈBЈ©»щУЪRT-qPCRөДMeRIPҷzңyNSUN2»щТтЗГіэөДҪӣ(jЁ©ng)»тОҙҪӣ(jЁ©ng)NSUN2ТЦЦЖ„©МҺАнөДCal-62јҡ°ыЦРSRSF6 mRNAөДm5Cё»јҜ·ЦОцЎЈ
CЈ©НЁЯ^qRT-PCRҢҚтһФu№АNSUN2»щТтЗГіэөДҪӣ(jЁ©ng)»тОҙҪӣ(jЁ©ng)NSUN2ТЦЦЖ„©МҺАнөДCal-62јҡ°ыЦРSRSF6 mRNAөДәЛЩ|(zhЁ¬)·ЦІјЎЈ
DЈ©НЁЯ^Western blot·ЦОцNSUN2»щТтЗГіэөДҪӣ(jЁ©ng)»тОҙҪӣ(jЁ©ng)NSUN2ТЦЦЖ„©МҺАнөДCal-62јҡ°ыЦРЦёКҫө°°ЧөДЛ®ЖҪЎЈ
EЈ©RT-PCRп@КҫҪӣ(jЁ©ng)»тОҙҪӣ(jЁ©ng)NSUN2ТЦЦЖ„©МҺАнөДCal-62јҡ°ыәНACT-1јҡ°ыЦРUAP1өДјфҪУКВјюЎЈ
FЈ©ҷzңyҝӮө°°ЧөДҝјсRЛ№ББЛ{ИҫЙ«ЈЁЧуЈ©Ј¬Ҫӣ(jЁ©ng)»тОҙҪӣ(jЁ©ng)NSUN2ТЦЦЖ„©МҺАнөДCal-62јҡ°ыәНACT-1јҡ°ыөДМЗө°°ЧИҫЙ«ЈЁУТЈ©ЎЈ
GЈ©НЁЯ^Western blot·ЦОцҪӣ(jЁ©ng)»тОҙҪӣ(jЁ©ng)NSUN2ТЦЦЖ„©МҺАнөДCal-62јҡ°ыәНACT-1јҡ°ыЦРЦёКҫө°°ЧөДЛ®ЖҪЎЈ
H-JЈ©НЁЯ^БчКҪјҡ°ығx·ЦОцҪӣ(jЁ©ng)»тОҙҪӣ(jЁ©ng)NSUN2ТЦЦЖ„©МҺАнөДCal-62јҡ°ыЦР¶аИбұИРЗЎўнҳгKәНҒц·ҘМжДбөДјҡ°ығИ(nЁЁi)·eАЫЗйӣrЎЈУТӮИ(cЁЁ)п@КҫЖҪҫщҹЙ№вҸҠ¶ИөД¶ЁБҝҪY(jiЁҰ)№ыЎЈ
K-MЈ©јҡ°ыЙъйLТЦЦЖҢҚтһФu№АNSUN2ТЦЦЖ„©Еc¶аИбұИРЗЎўнҳгK»тҒц·ҘМжДбВ“(liЁўn)әПК№УГҢҰCal-62јҡ°ыөДУ°н‘ЎЈ
NЈ©Х№КҫФЪ»щТт№ӨіМРЎКуЦРNSUN2ТЦЦЖ„©Ўў¶аИбұИРЗЎўнҳгKәНҒц·ҘМжДбөДҪoЛҺ·Ҫ°ёКҫТвҲDЎЈ
OЈ©ЙўьcҲDп@КҫёчҪMРЎКуөДЧоҪKД[БцЦШБҝЎЈ
PЈ©ЙўьcҲDп@КҫёчҪMРЎКуөДЧоҪKД[Бцуw·eЎЈ
QЈ©п@КҫёчҪMATCРЎКуөДЙъҙжЗъҫҖЎЈ
ҲD9ЈәNSUN2ФЪATC¶аЛҺДНЛҺЦРөД№ҰДЬәНҷCЦЖКҫТвҲDДЈРНЎЈ
У‘Х“әНҫЦПЮРФ
ұҫСРҫҝҪТКҫБЛNSUN2НЁЯ^m5CРЮп—Х{(diЁӨo)№қ(jiЁҰ)SRSF6өДјҡ°ыәЛЩ|(zhЁ¬)ЮD(zhuЁЈn)Я\Ј¬ЯM¶шУ°н‘UAP1јфЗРәНN-МЗ»щ»ҜЛ®ЖҪЈ¬ЧоҪKҢ§(dЁЈo)ЦВATCДНЛҺРФөДҷCЦЖЎЈСРҫҝҪY(jiЁҰ)№ыұнГчЈ¬NSUN2ј°ЖдПВУО°РьcКЗ°РПтATCДНЛҺРФөДқ“ФЪЦОҜҹ°РьcЎЈ
И»¶шЈ¬NSUN2ФЪХэіЈјҡ°ыЙъАнЦРТІ°l(fЁЎ)“]ЦШТӘЧчУГЈ¬ПөҪy(tЁҜng)РФТЦЦЖNSUN2ҝЙДЬ•юҺ§Ғн°ІИ«РФҶ–о}ЎЈҙЛНвЈ¬ИзәОҢҚ¬F(xiЁӨn)NSUN2ТЦЦЖ„©өДД[БцМШ®җРФЯfЛНИФИ»КЗТ»ӮҖМф‘р(zhЁӨn)ЎЈ
ТЧРЎҪY(jiЁҰ)
ұҫСРҫҝНЁЯ^m5C MeRIP-seqәНRNA-seqөИ·ЦОцҪТКҫБЛNSUN2/SRSF6/UAP1РЕМ–ЭSФЪATCДНЛҺРФЦРөДЧчУГҷCЦЖЈ¬һйй_°l(fЁЎ)РВөДЦОҜҹІЯВФМṩБЛАнХ“ТА“ю(jЁҙ)ЎЈNSUN2ТЦЦЖ„©Еc¬F(xiЁӨn)УРҝ№°©ЛҺОпөДВ“(liЁўn)әПК№УГУРНыіЙһйҝЛ·юATCДНЛҺРФөДРВ·Ҫ·ЁЎЈОҙҒнөДСРҫҝРиТӘЯMТ»ІҪФu№АNSUN2ТЦЦЖ„©өД°ІИ«РФәНУРР§РФЈ¬ІўМҪЛчЖдФЪЖдЛыД[БцоҗРНЦРөДқ“ФЪ‘Ә(yЁ©ng)УГЎЈ
m5C MeRIP-seqФЪұҫСРҫҝЦРөДЦШТӘЧчУГ
ұҫСРҫҝөДm5C MeRIP-seqЈЁјЧ»щ»ҜRNAГвТЯіБөнңyРтЈ©ЦчТӘУГУЪҪТКҫNSUN2ФЪјЧ оПЩОҙ·Ц»Ҝ°©ЈЁATCЈ©ЦРөДЧчУГҷCЦЖЈ¬ЦчТӘұн¬F(xiЁӨn)ФЪЈә
ЈЁ1Ј©иb¶Ёm5CРЮп—өД°Р»щТтЈәНЁЯ^MeRIP-seqјјРg(shЁҙ)ДЬүтПөҪy(tЁҜng)өШ·ЦОцNSUN2ФЪATCјҡ°ыЦРҢҰmRNAөДm5CРЮп—ЗйӣrЎЈСРҫҝ°l(fЁЎ)¬F(xiЁӨn)Ј¬NSUN2ДЬүтМШ®җРФөШРЮп—SRSF6 mRNAЙПөДm5CО»ьcЎЈЯ@Т»°l(fЁЎ)¬F(xiЁӨn)һйәуАm(xЁҙ)СРҫҝNSUN2ИзәОНЁЯ^m5CРЮп—Х{(diЁӨo)ҝШ»щТтұнЯ_МṩБЛЦұҪУЧC“ю(jЁҙ)ЎЈ
ЈЁ2Ј©ҪТКҫm5CРЮп—өД№ҰДЬЈәMeRIP-seq·ЦОцп@КҫЈ¬NSUN2ҪйҢ§(dЁЈo)өДm5CРЮп—ДЬүтХ{(diЁӨo)№қ(jiЁҰ)SRSF6 mRNAөДјҡ°ыәЛЩ|(zhЁ¬)ЮD(zhuЁЈn)Я\ЎЈҫЯуwҒнХfЈ¬m5CРЮп—ФцҸҠБЛSRSF6 mRNAҸДјҡ°ыәЛПтјҡ°ыЩ|(zhЁ¬)өДЮD(zhuЁЈn)Я\Ј¬ҸД¶шҙЩЯMБЛSRSF6ө°°ЧөДұнЯ_ЎЈЯ@Т»Я^іМҢҰУЪATCјҡ°ыөД¶аЛҺДНЛҺРФЈЁMDRЈ©ЦБкP(guЁЎn)ЦШТӘЎЈ
ЈЁ3Ј©МҪЛчm5CРЮп—өДХ{(diЁӨo)ҝШҷCЦЖЈәСРҫҝЦРНЁЯ^MeRIP-seqҪY(jiЁҰ)әПЖдЛыҢҚтһКЦ¶ОЈЁИзRNAПВАӯҢҚтһәНRIP-qPCRЈ©Ј¬ҪТКҫБЛALYREFЧчһйm5CРЮп—өД“reader”Ј¬ДЬүтиb¶ЁІўҪY(jiЁҰ)әПm5CРЮп—өДSRSF6 mRNAЈ¬ІўҢўЖдҸДјҡ°ыәЛЮD(zhuЁЈn)Я\өҪјҡ°ыЩ|(zhЁ¬)ЎЈЯ@Т»°l(fЁЎ)¬F(xiЁӨn)ЯMТ»ІҪНкЙЖБЛm5CРЮп—ФЪ»щТтұнЯ_Х{(diЁӨo)ҝШЦРөД·ЦЧУҷCЦЖЎЈ
ЈЁ4Ј©тһЧCm5CРЮп—өДЙъОпҢW(xuЁҰ)ТвБxЈәНЁЯ^ұИЭ^NSUN2ЗГіэјҡ°ыЕcТ°ЙъРНјҡ°ыөДMeRIP-seq”ө(shЁҙ)“ю(jЁҙ)Ј¬СРҫҝИЛҶT°l(fЁЎ)¬F(xiЁӨn)NSUN2ЗГіэәуSRSF6 mRNAөДm5CРЮп—Л®ЖҪп@ЦшПВҪөЈ¬Н¬•rSRSF6ө°°ЧұнЯ_ТІКЬөҪТЦЦЖЎЈЯ@ұнГчm5CРЮп—ФЪҫSіЦSRSF6№ҰДЬәНATCјҡ°ыДНЛҺРФЦР°l(fЁЎ)“]ЦшЦШТӘЧчУГЎЈ
…ўҝјОД«IЈә
Hou X, Dong Q, Hao J, Liu M, Ning J, Tao M, Wang Z, Guo F, Huang D, Shi X, Gao M, Li D, Zheng X. NSUN2-mediated m5C modification drives alternative splicing reprogramming and promotes multidrug resistance in anaplastic thyroid cancer through the NSUN2/SRSF6/UAP1 signaling axis. Theranostics 2025; 15(7):2757-2777. doi:10.7150/thno.104713.
ҳЛ(biЁЎo)әһЈә
RNAјЧ»щ»Ҝ

- Target-BSјјРg(shЁҙ)ҪТКҫМЗДтІЎТэ°l(fЁЎ)ІӘЖр№ҰДЬХПөKөДDNAјЧ»щ»ҜХ{(diЁӨo)ҝШҷCЦЖ
- әПіЙЙъОпҢW(xuЁҰ)өЧұPјҡ°ыЦ®®…іаҪНДёіЈУГұнЯ_Эdуwј°»щТтёДФмјјРg(shЁҙ)
- Йо¶ИҪвОцCRISPRОДҺмәYЯxБчіМј°‘Ә(yЁ©ng)УГ°ёАэ
- ҝЙңШәН—lјюПВёЯ·ЦұжВКҷzңym6AРЮп—өДРВm6AңyРт·Ҫ·Ёй_°l(fЁЎ)СРҫҝ
- AIИЛ№ӨЦЗДЬФЪө°°ЧЩ|(zhЁ¬)ҪY(jiЁҰ)ҳӢ(gЁ°u)оA(yЁҙ)ңyЎў№ҰДЬоA(yЁҙ)ңyј°ФO(shЁЁ)УӢЦРөДЧчУГ
- m5C MeRIP-seqөИҪТКҫm5CРЮп—ФЪ°©°YДНЛҺЦРөДкP(guЁЎn)жIХ{(diЁӨo)ҝШҷCЦЖЦРөД‘Ә(yЁ©ng)УГ
- ChIPјјРg(shЁҙ)ҪТКҫNURR1ФЪЗ°БРПЩ°©ҸД»щТтЮD(zhuЁЈn)дӣөҪД[БцЯMХ№ЦРөДХ{(diЁӨo)ҝШҷCЦЖ
- ғЙ·NЮD(zhuЁЈn)дӣТтЧУФЪҪйҢ§(dЁЈo)·¬ЗС№ыҢҚіЙКмөДұнУ^ЯzӮчХ{(diЁӨo)ҝШЦР°l(fЁЎ)“]өДкP(guЁЎn)жIЧчУГ
- 2025І®әАЙъОпҙәјҫИ«ҮшСІЦvй_ҶўЈ¬ФзшBҲуГы“ҢХјПИҷC
- ИAҙуЦЗФмёұҝӮІГЦРҮш…^(qЁұ)ҝӮҪӣ(jЁ©ng)АнЕнҡgҡgТ»РРөҪФLІ®әАЙъОп
- І®әА¶ӯКВйLіцПҜҢЈҫ«МШРВЦРРЎЖуҳI(yЁЁ)°l(fЁЎ)Х№үСҙуХ“үҜІў°l(fЁЎ)СФ
- І®әАЙъОплp11ҝсҡgМШ»Э»о„УҒнТuЈ¬ПЮ•rПЮБҝ“Ң
- вщГАНЁөВНЖіціЙұҫғrуwтһҶОјҡ°ыңyРтЈЁFFPE»тИ«СӘЈ©
- І®әАЙъОп2024ПөБРЕаУ–(xЁҙn)°аЈЁкғОчХҫЈ©ҲуГый_Ҷў
- І®әАҶОјҡ°ыFlexДкЦРҫЮ»ЭЈ¬1.1W/ҳУұҫ»о„УғrјҙҢўҪШЦ№
- Л{ҫ°ҝЖРЕDAP-seqјјРg(shЁҙ)ПакP(guЁЎn)ОДХВ6ЯB°l(fЁЎ)Ј¬ҝӮIF 95.2



Copyright(C) 1998-2025 ЙъОпЖчІДҫW(wЁЈng) лҠФ’Јә021-64166852;13621656896 E-mailЈәinfo@bio-equip.com