
底物特異性和蛋白質穩定性促進植物特異性DNA甲基轉移酶分化
大家好,這里是專注表觀組學十余年,領跑多組學科研服務的易基因。
DNA甲基化是一種重要的表觀遺傳機制,對轉座子沉默和基因組完整性至關重要。在進化過程中,DNA甲基化底物在不同生物界中發生多樣化。在植物中,染色體甲基化酶3(CMT3)和CMT2分別介導CHG和CHH位點的甲基化。然而,這兩種甲基轉移酶在進化過程中如何分化其底物特異性仍然未知。
近日,河南大學蔣建軍教授為第一作者,美國圣路易斯華盛頓大學鐘雪花教授和加州大學河濱分校宋吉奎教授為共同通訊在Science子刊《Science Advances》上發表了題為“Substrate specificity and protein stability drive the divergence of plant-specific DNA methyltransferases”的研究論文,揭示了底物特異性和蛋白質穩定性如何驅動植物特異性DNA甲基轉移酶分化,特別是CMT2和CMT3在進化過程中的底物特異性分化,闡明了CMT2和CMT3產生功能分化的表觀遺傳調控機制。

標題:Substrate specificity and protein stability drive the divergence of plant-specific DNA methyltransferases(底物特異性和蛋白質穩定性促進植物特異性DNA甲基轉移酶分化)
發表時間:2024年11月6日
發表期刊:Science Advances
影響因子:IF 11.7 / 1區
應用技術:BS-seq、RNA-seq(易基因金牌技術)
本研究進行了全面的結構、功能和進化研究,分析了CMT2和CMT3分化背后的分子機制。研究首先揭示了對CMT3識別CHG至關重要的精氨酸殘基(R745)在CMT2中表現出很大的變異,解釋了其失去CHG特異性的原因。在擬南芥中,將CMT2中相應的殘基突變為精氨酸(V1200R)獲得了CHG甲基化活性,并在cmt2cmt3突變體中重新沉默一組轉座元件(TEs),發揮了類似CMT3功能。CMT3有一個短的N末端,CMT2卻包含一個長而無序的N末端,這是許多植物物種的共同特征。這個長的N末端調控CMT2穩定性,并介導熱誘導的CMT2降解。此外由于自然界中CMT2的N端存在多種變異,CMT2的N端更具可塑性,能夠高度容忍各種自然變異。總的來說,本研究揭示了植物種染色體甲基化酶靶向特定環境DNA甲基化的分化機制,并為植物中DNA甲基化的功能和進化提供了新的視角。
研究方法:
1) 結構和功能研究:通過比較CMT3和CMT2的關鍵殘基,構建了CMT2與DNA復合物的結構模型,并進行體外甲基化實驗。
2) 系統發育分析:通過系統發育分析探討CMT2和CMT3的進化關系。
3) 基因編輯:通過在擬南芥中引入V1200R突變,研究者們增強了CMT2的CHG甲基化活性。
4) DNA甲基化分析:利用全基因組亞硫酸鹽測序(WGBS-seq)技術分析CMT2和CMT2V1200R對全基因組甲基化水平的影響。
5) 轉錄組測序:通過RNA-seq分析CMT2V1200R介導CHG甲基化對基因表達的影響。
6) 自然變異分析:分析自然條件下的CMT2變異,以及這些變異對DNA甲基化和植物適應性影響。
結果圖形
(1)CMT2起源于開花植物中CMT3的復制,CMT2V1200R在體外和體內誘導CHG甲基化增加
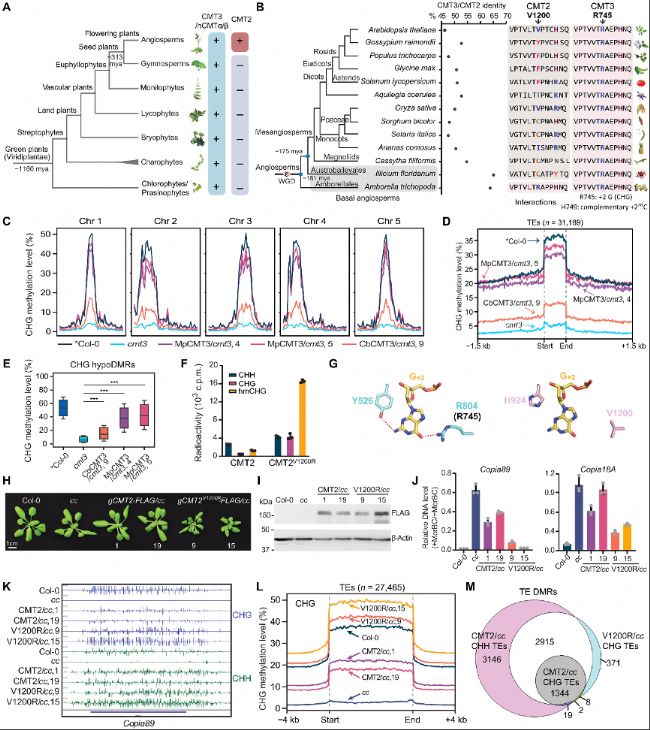
圖1:精氨酸殘基丟失使CMT2具有與CMT3不同的底物偏好。
A. 綠色植物(Viridiplantae)中CMT3(或其近緣同源物hCMTα/β)和CMT2的系統發育樹。
B. CMT2和CMT3之間的蛋白質序列同一性(左圖)以及在開花植物中與+2鳥嘌呤(G+2)接觸的殘基和CMT2中的相應殘基(右圖)。
C. 在cmt3突變背景下,M. polymorpha CMT3(MpCMT3)和C. braunii CMT3(CbCMT3)的CHG甲基化水平Meta圖。
D. 擬南芥中所有轉座元件(TEs)上的平均CHG甲基化水平Meta圖。
E. CHG低甲基化區域(hypoDMRs)上的CHG甲基化水平箱線圖。
F. CMT2和CMT2V1200R對CHH、CHG或半甲基化CHG(hmCHG)DNA底物的體外DNA甲基轉移酶檢測。
G. 玉米ZMET2-DNA復合物結構(蛋白質數據庫7UBU)顯示ZMET2 Y526和R804與CHG環境中目標胞嘧啶+2位點(G+2)鳥嘌呤之間的相互作用。
H. 兩個獨立的轉基因CMT2和CMT2V1200R植物在cmt2cmt3(cc)突變背景下的表型圖像。
I. 免疫印跡圖顯示(H)中植物的CMT2和CMT2V1200R蛋白水平。
J. McrBC-qPCR分析兩個CMT2靶向TEs Copia89和Copia18A的DNA甲基化水平。
K. BS-seq中Copia89的CHG和CHH甲基化水平的基因組瀏覽器視圖。
L. 擬南芥中所有TEs上的平均CHG甲基化水平Meta圖。
M. 與Col-0比較時鑒定DMRs的重疊情況維恩圖。
(2)CMT2V1200R介導的CHG甲基化抑制TE
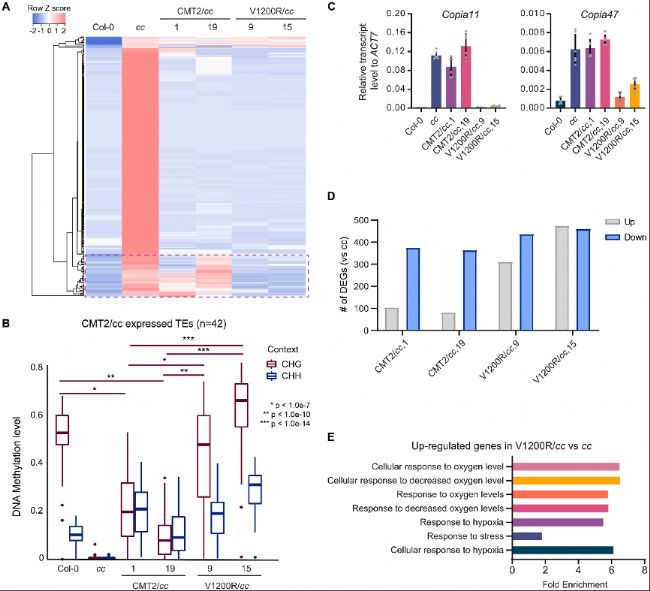
圖2. CMT2V1200R抑制CMT2和CMT3靶向的轉座元件(TEs)。
A. 兩個獨立的CMT2/cc和V1200R/cc轉基因系中上調TEs的表達水平熱圖。cc代表cmt2cmt3。虛線框表示被CMT2V1200R重新沉默的TEs。
B. CMT2/cc中仍然表達的TEs的CHG和CHH DNA甲基化水平。
C. 在CMT2/cc和V1200R/cc轉基因植物中,通過RT-qPCR檢測Copia11和Copia47的相對轉錄水平。
D. CMT2/cc和V1200R/cc中差異表達基因(DEGs)數量。
E. V1200R/cc中上調基因的基因本體(GO)分析。
(3)CMT2包含一個對核定位很重要的長N末端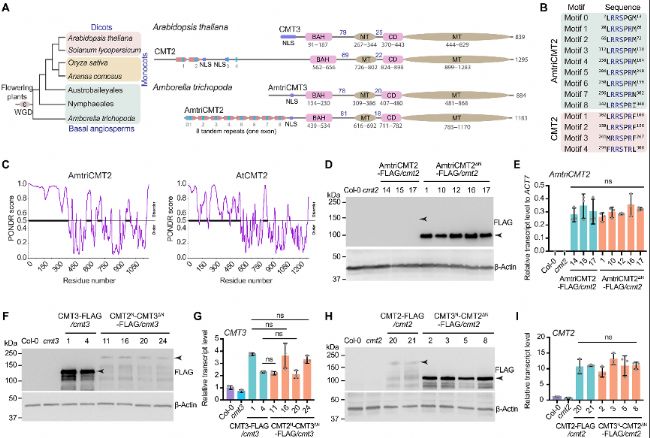
B. A.trichopoda和A. thaliana CMT2 N端保守RRS motif比對。
C. A.trichopoda和A. thaliana CMT2的固有無序得分圖,由PONDR評分。
D. 在A. thaliana cmt2突變背景下表達全長A.trichopoda CMT2(AmtriCMT2)和N端截短的A.trichopoda CMT2(AmtriCMT2ΔN)的蛋白水平免疫印跡。
E. RT-qPCR顯示AmtriCMT2和AmtriCMT2ΔN轉基因系中AmtriCMT2轉錄水平。
F. 野生型(CMT3)和N端轉換的CMT3(CMT2N-CMT3ΔN)轉基因系在cmt3突變體中的蛋白水平免疫印跡。
G. RT-qPCR顯示CMT3和CMT2N-CMT3ΔN轉基因植物中CMT3轉錄水平。
H. 野生型(CMT2)和N端轉換的CMT2(CMT3N-CMT2ΔN)轉基因系在A. thaliana cmt2突變體中的蛋白水平免疫印跡。
I. RT-qPCR顯示CMT2和CMT3N-CMT2ΔN轉基因系中CMT2轉錄水平。
轉錄水平首先歸一化到Col-0中的CMT2水平,然后歸一化到Col-0中的ACT7水平。ns,無顯著性差異。
(4)CMT2的N末端調控蛋白質穩定性
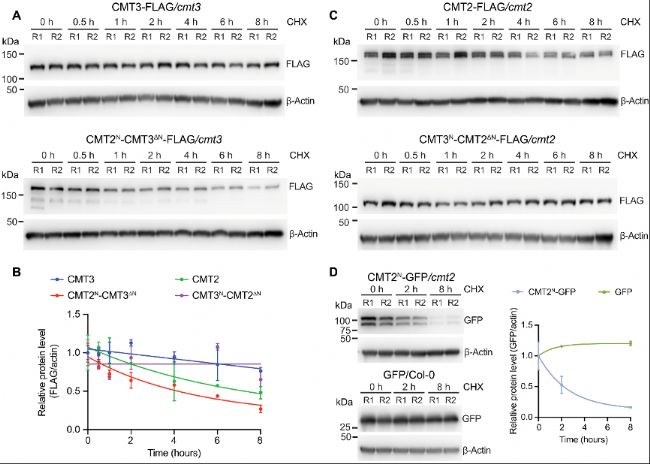
B. 相對于肌動蛋白歸一化的蛋白水平定量。
C. 免疫印跡圖顯示用500 μM環己酰亞胺處理指定時間后CMT2(頂部)和CMT3N-CMT2ΔN(底部)的水平。
D. 免疫印跡圖顯示用500 μM環己酰亞胺處理后CMT2N-GFP(頂部)和僅GFP(底部)的水平。右側是通過GFP信號相對于肌動蛋白標準化的蛋白水平定量。
(5)N末端介導熱誘導的CMT2降解

圖5. 長N末端介導熱誘導的CMT2蛋白降解。
A. 免疫印跡圖顯示在37°C熱處理指定時間后CMT3(頂部)和CMT2N-CMT3ΔN(底部)的水平。使用10日齡幼苗。
B. 相對于肌動蛋白歸一化的蛋白水平定量。
C. 免疫印跡圖顯示在37°C熱處理指定時間后CMT2(頂部)和CMT3N-CMT2ΔN(底部)的水平。
D. 免疫印跡圖顯示在37°C熱處理后CMT2N-GFP(頂部)和僅GFP(底部)的水平。
E. 共聚焦顯微鏡圖像顯示在37°C熱處理指定時間后,擬南芥根尖中CMT2N-GFP和CMT2-GFP的定位和強度。
(6)CMT2自然變異表現出對環境脅迫的耐受性
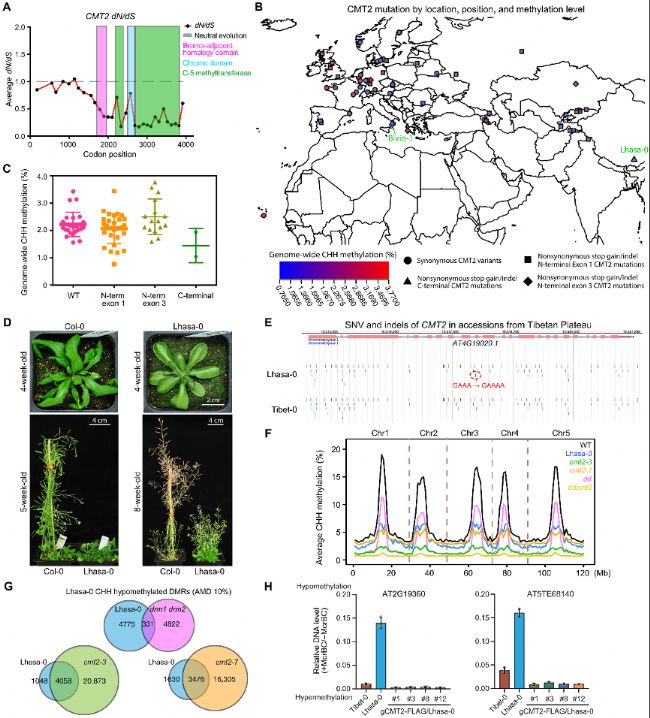
B. 不同CMT2基因突變的品系全基因組CHH甲基化水平。
C. 野生型(WT;n=25)、N端(第1外顯子(n=31)和第3外顯子(n=16)的AT4G19020.1)和C端(n=2)CMT2突變的CHH甲基化點圖。
D. 擬南芥生態型Col-0和Lhasa-0的表型比較。
E. 來自青藏高原的自然品系Lhasa-0和Tibet-0的基因組瀏覽器視圖,顯示CMT2外顯子中的單核苷酸變異(SNV)和插入。
F. 與野生型Col-0和CHH DNA甲基轉移酶突變體相比,Lhasa-0生態型在擬南芥染色體上的平均水平CHH甲基化。
G. Lhasa-0與CHH DNA甲基轉移酶突變體之間低甲基化差異甲基化區域(DMRs)的重疊。
H. McrBC-qPCR分析顯示在兩個代表性位點的DNA甲基化互補。含有CMT2基因組序列(gCMT2)的轉基因系可以挽救Lhasa-0中丟失的CHH甲基化。Tibet-0用作對照。AMD,絕對甲基化差異。
易小結
本研究通過WGBS+RNA-seq等分析揭示了植物中CMTs的分化機制,為理解DNA甲基化在植物進化和功能中的作用提供了重要見解。
研究亮點
1) CMT2的起源和進化:研究揭示了CMT2是如何從CMT3分化而來,以及這一分化如何影響了植物的DNA甲基化模式。
2) V1200R突變的功能研究:通過V1200R突變,研究者們成功地改變了CMT2的底物特異性,這一發現對于理解酶活性的調控具有重要意義。
3) CMT2 N端的穩定性研究:研究者們發現CMT2的長N末端在熱應激下對其穩定性至關重要,這一發現為理解環境因素如何影響表觀遺傳機制提供了新的視角。
4) 自然變異與環境適應性:通過分析自然條件下CMT2的變異,研究者們揭示了這些變異如何影響植物對環境應激的適應性,這對于理解植物進化和育種具有重要意義。
這項研究不僅增進了對植物DNA甲基化機制的理解,還為農業育種和作物改良提供了潛在的分子靶標。
參考文獻:
Jiang J, Gwee J, Fang J, Leichter SM, Sanders D, Ji X, Song J, Zhong X. Substrate specificity and protein stability drive the divergence of plant-specific DNA methyltransferases. Sci Adv. 2024 Nov 8;10(45):eadr2222. doi: 10.1126/sciadv.adr2222. PubMed PMID: 39504374.
DNA甲基化是一種重要的表觀遺傳機制,對轉座子沉默和基因組完整性至關重要。在進化過程中,DNA甲基化底物在不同生物界中發生多樣化。在植物中,染色體甲基化酶3(CMT3)和CMT2分別介導CHG和CHH位點的甲基化。然而,這兩種甲基轉移酶在進化過程中如何分化其底物特異性仍然未知。
近日,河南大學蔣建軍教授為第一作者,美國圣路易斯華盛頓大學鐘雪花教授和加州大學河濱分校宋吉奎教授為共同通訊在Science子刊《Science Advances》上發表了題為“Substrate specificity and protein stability drive the divergence of plant-specific DNA methyltransferases”的研究論文,揭示了底物特異性和蛋白質穩定性如何驅動植物特異性DNA甲基轉移酶分化,特別是CMT2和CMT3在進化過程中的底物特異性分化,闡明了CMT2和CMT3產生功能分化的表觀遺傳調控機制。
標題:Substrate specificity and protein stability drive the divergence of plant-specific DNA methyltransferases(底物特異性和蛋白質穩定性促進植物特異性DNA甲基轉移酶分化)
發表時間:2024年11月6日
發表期刊:Science Advances
影響因子:IF 11.7 / 1區
應用技術:BS-seq、RNA-seq(易基因金牌技術)
本研究進行了全面的結構、功能和進化研究,分析了CMT2和CMT3分化背后的分子機制。研究首先揭示了對CMT3識別CHG至關重要的精氨酸殘基(R745)在CMT2中表現出很大的變異,解釋了其失去CHG特異性的原因。在擬南芥中,將CMT2中相應的殘基突變為精氨酸(V1200R)獲得了CHG甲基化活性,并在cmt2cmt3突變體中重新沉默一組轉座元件(TEs),發揮了類似CMT3功能。CMT3有一個短的N末端,CMT2卻包含一個長而無序的N末端,這是許多植物物種的共同特征。這個長的N末端調控CMT2穩定性,并介導熱誘導的CMT2降解。此外由于自然界中CMT2的N端存在多種變異,CMT2的N端更具可塑性,能夠高度容忍各種自然變異。總的來說,本研究揭示了植物種染色體甲基化酶靶向特定環境DNA甲基化的分化機制,并為植物中DNA甲基化的功能和進化提供了新的視角。
研究方法:
1) 結構和功能研究:通過比較CMT3和CMT2的關鍵殘基,構建了CMT2與DNA復合物的結構模型,并進行體外甲基化實驗。
2) 系統發育分析:通過系統發育分析探討CMT2和CMT3的進化關系。
3) 基因編輯:通過在擬南芥中引入V1200R突變,研究者們增強了CMT2的CHG甲基化活性。
4) DNA甲基化分析:利用全基因組亞硫酸鹽測序(WGBS-seq)技術分析CMT2和CMT2V1200R對全基因組甲基化水平的影響。
5) 轉錄組測序:通過RNA-seq分析CMT2V1200R介導CHG甲基化對基因表達的影響。
6) 自然變異分析:分析自然條件下的CMT2變異,以及這些變異對DNA甲基化和植物適應性影響。
結果圖形
(1)CMT2起源于開花植物中CMT3的復制,CMT2V1200R在體外和體內誘導CHG甲基化增加
圖1:精氨酸殘基丟失使CMT2具有與CMT3不同的底物偏好。
B. CMT2和CMT3之間的蛋白質序列同一性(左圖)以及在開花植物中與+2鳥嘌呤(G+2)接觸的殘基和CMT2中的相應殘基(右圖)。
C. 在cmt3突變背景下,M. polymorpha CMT3(MpCMT3)和C. braunii CMT3(CbCMT3)的CHG甲基化水平Meta圖。
D. 擬南芥中所有轉座元件(TEs)上的平均CHG甲基化水平Meta圖。
E. CHG低甲基化區域(hypoDMRs)上的CHG甲基化水平箱線圖。
F. CMT2和CMT2V1200R對CHH、CHG或半甲基化CHG(hmCHG)DNA底物的體外DNA甲基轉移酶檢測。
G. 玉米ZMET2-DNA復合物結構(蛋白質數據庫7UBU)顯示ZMET2 Y526和R804與CHG環境中目標胞嘧啶+2位點(G+2)鳥嘌呤之間的相互作用。
H. 兩個獨立的轉基因CMT2和CMT2V1200R植物在cmt2cmt3(cc)突變背景下的表型圖像。
I. 免疫印跡圖顯示(H)中植物的CMT2和CMT2V1200R蛋白水平。
J. McrBC-qPCR分析兩個CMT2靶向TEs Copia89和Copia18A的DNA甲基化水平。
K. BS-seq中Copia89的CHG和CHH甲基化水平的基因組瀏覽器視圖。
L. 擬南芥中所有TEs上的平均CHG甲基化水平Meta圖。
M. 與Col-0比較時鑒定DMRs的重疊情況維恩圖。
(2)CMT2V1200R介導的CHG甲基化抑制TE
圖2. CMT2V1200R抑制CMT2和CMT3靶向的轉座元件(TEs)。
B. CMT2/cc中仍然表達的TEs的CHG和CHH DNA甲基化水平。
C. 在CMT2/cc和V1200R/cc轉基因植物中,通過RT-qPCR檢測Copia11和Copia47的相對轉錄水平。
D. CMT2/cc和V1200R/cc中差異表達基因(DEGs)數量。
E. V1200R/cc中上調基因的基因本體(GO)分析。
(3)CMT2包含一個對核定位很重要的長N末端
圖3. CMT2的長N端無序,且調控蛋白水平。
A. 擬南芥(A. thaliana,雙子葉植物)和A.trichopoda(基部被子植物)的CMT2和CMT3域結構圖表。藍色條表示串聯重復。MT,甲基轉移酶域;CD,染色質域。線上的數字表示域之間的氨基酸數量。WGD,全基因組復制。B. A.trichopoda和A. thaliana CMT2 N端保守RRS motif比對。
C. A.trichopoda和A. thaliana CMT2的固有無序得分圖,由PONDR評分。
D. 在A. thaliana cmt2突變背景下表達全長A.trichopoda CMT2(AmtriCMT2)和N端截短的A.trichopoda CMT2(AmtriCMT2ΔN)的蛋白水平免疫印跡。
E. RT-qPCR顯示AmtriCMT2和AmtriCMT2ΔN轉基因系中AmtriCMT2轉錄水平。
F. 野生型(CMT3)和N端轉換的CMT3(CMT2N-CMT3ΔN)轉基因系在cmt3突變體中的蛋白水平免疫印跡。
G. RT-qPCR顯示CMT3和CMT2N-CMT3ΔN轉基因植物中CMT3轉錄水平。
H. 野生型(CMT2)和N端轉換的CMT2(CMT3N-CMT2ΔN)轉基因系在A. thaliana cmt2突變體中的蛋白水平免疫印跡。
I. RT-qPCR顯示CMT2和CMT3N-CMT2ΔN轉基因系中CMT2轉錄水平。
轉錄水平首先歸一化到Col-0中的CMT2水平,然后歸一化到Col-0中的ACT7水平。ns,無顯著性差異。
(4)CMT2的N末端調控蛋白質穩定性
圖4. CMT2的N末端調控其蛋白穩定性。
A. 用500 μM環己酰亞胺(CHX)處理指定時間后CMT3(頂部)和CMT2N-CMT3ΔN(底部)的水平免疫印跡圖。使用7日齡幼苗。肌動蛋白作為對照。R1和R2代表兩個生物學重復。B. 相對于肌動蛋白歸一化的蛋白水平定量。
C. 免疫印跡圖顯示用500 μM環己酰亞胺處理指定時間后CMT2(頂部)和CMT3N-CMT2ΔN(底部)的水平。
D. 免疫印跡圖顯示用500 μM環己酰亞胺處理后CMT2N-GFP(頂部)和僅GFP(底部)的水平。右側是通過GFP信號相對于肌動蛋白標準化的蛋白水平定量。
(5)N末端介導熱誘導的CMT2降解
圖5. 長N末端介導熱誘導的CMT2蛋白降解。
B. 相對于肌動蛋白歸一化的蛋白水平定量。
C. 免疫印跡圖顯示在37°C熱處理指定時間后CMT2(頂部)和CMT3N-CMT2ΔN(底部)的水平。
D. 免疫印跡圖顯示在37°C熱處理后CMT2N-GFP(頂部)和僅GFP(底部)的水平。
E. 共聚焦顯微鏡圖像顯示在37°C熱處理指定時間后,擬南芥根尖中CMT2N-GFP和CMT2-GFP的定位和強度。
(6)CMT2自然變異表現出對環境脅迫的耐受性
圖6. 擬南芥中CMT2的自然變異。
A. 擬南芥近緣種CMT2基因的dN/dS繪圖。B. 不同CMT2基因突變的品系全基因組CHH甲基化水平。
C. 野生型(WT;n=25)、N端(第1外顯子(n=31)和第3外顯子(n=16)的AT4G19020.1)和C端(n=2)CMT2突變的CHH甲基化點圖。
D. 擬南芥生態型Col-0和Lhasa-0的表型比較。
E. 來自青藏高原的自然品系Lhasa-0和Tibet-0的基因組瀏覽器視圖,顯示CMT2外顯子中的單核苷酸變異(SNV)和插入。
F. 與野生型Col-0和CHH DNA甲基轉移酶突變體相比,Lhasa-0生態型在擬南芥染色體上的平均水平CHH甲基化。
G. Lhasa-0與CHH DNA甲基轉移酶突變體之間低甲基化差異甲基化區域(DMRs)的重疊。
H. McrBC-qPCR分析顯示在兩個代表性位點的DNA甲基化互補。含有CMT2基因組序列(gCMT2)的轉基因系可以挽救Lhasa-0中丟失的CHH甲基化。Tibet-0用作對照。AMD,絕對甲基化差異。
易小結
本研究通過WGBS+RNA-seq等分析揭示了植物中CMTs的分化機制,為理解DNA甲基化在植物進化和功能中的作用提供了重要見解。
研究亮點
1) CMT2的起源和進化:研究揭示了CMT2是如何從CMT3分化而來,以及這一分化如何影響了植物的DNA甲基化模式。
2) V1200R突變的功能研究:通過V1200R突變,研究者們成功地改變了CMT2的底物特異性,這一發現對于理解酶活性的調控具有重要意義。
3) CMT2 N端的穩定性研究:研究者們發現CMT2的長N末端在熱應激下對其穩定性至關重要,這一發現為理解環境因素如何影響表觀遺傳機制提供了新的視角。
4) 自然變異與環境適應性:通過分析自然條件下CMT2的變異,研究者們揭示了這些變異如何影響植物對環境應激的適應性,這對于理解植物進化和育種具有重要意義。
這項研究不僅增進了對植物DNA甲基化機制的理解,還為農業育種和作物改良提供了潛在的分子靶標。
參考文獻:
Jiang J, Gwee J, Fang J, Leichter SM, Sanders D, Ji X, Song J, Zhong X. Substrate specificity and protein stability drive the divergence of plant-specific DNA methyltransferases. Sci Adv. 2024 Nov 8;10(45):eadr2222. doi: 10.1126/sciadv.adr2222. PubMed PMID: 39504374.
標簽:
DNA甲基化




Copyright(C) 1998-2025 生物器材網 電話:021-64166852;13621656896 E-mail:info@bio-equip.com