
準確解讀細胞死亡實驗結果的技巧分享
摘要:細胞死亡是一個重要的生物過程,其檢測和測量過程困難,尤其是在多種檢測條件同時進行時。由于測量技術的復雜性和該領域標準化方法的缺乏,細胞死亡數據的解讀變得更加復雜。本文提供更多提示來幫助解釋細胞死亡實驗。
關鍵詞:細胞死亡、數據解讀、細胞增殖、細胞死亡機制、細胞分析、細胞生物學、細胞計數、活細胞成像
《Nature Cell Biology》期刊在2023年11月發表了一篇名為《Quick tips for interpreting cell death experiments》[1]的文章,文中列舉了解讀細胞死亡實驗結果的多個技巧。
細胞死亡是動物發育和維持體內穩態至關重要的環節,許多研究試圖了解細胞死亡是如何被調節的,也有研究試圖找到能夠觸發或防止細胞死亡的藥物。測量細胞死亡并清楚地呈現細胞死亡過程是一個艱巨的挑戰。這篇綜述提供了一些技巧來幫助解釋已發表的細胞死亡實驗的結果。重點在于培養細胞中進行的機制實驗,在兩種或兩種以上的條件下比較細胞死亡的數量,這是許多研究中常見的實驗方式。這些技巧是根據作者自己的研究、所遇到的問題和失誤,以及盡可能仔細地測量細胞死亡而做出的努力得出的。
Tip1:如何檢測細胞死亡
活細胞、瀕死細胞和死細胞都會產生許多信號,通過這些信號可直接檢測或間接推斷細胞死亡(圖1a,b),重要的是要注意不同技術是如何檢測細胞死亡的。例如,使用流式細胞術或圖像法直接對群體中的單個活細胞和/或死細胞進行計數。然而,許多研究中用來推斷細胞死亡的依據是群體中所有細胞獲得的大量代謝物,例如還原四唑基化合物(如使用MTT試劑盒)或測量總ATP豐度(如使用CellTiter-Glo)。雖然代謝數據相對簡單且易于拓展到大量樣品,但這些方法不能直接測量細胞死亡且會受到技術干擾(例如在細胞凋亡早期,線粒體損傷會改變新陳代謝、增加ATP水平,與細胞死亡無關[2]),在進行這些操作時,減緩新陳代謝但不殺死細胞的情況也可能引起誤解。監測細胞死亡途徑特異性生化變化(如凋亡過程中線粒體內膜跨膜電位的喪失)可能有助于分析細胞死亡機制的誘發情況,但仍會受到其他限制,下文將進行解析。
Tip2:如何分析細胞死亡
細胞死亡研究的常見比較方法是陰性對照與一種或多種處理條件之間進行對比(圖1c)。如上文所述,需要注意的是監測細胞死亡還是推斷細胞死亡。在使用大量代謝物進行檢測時,無法辨別治療處理是否會導致細胞死亡、減緩細胞增殖或導致這些效應的某種組合情況發生(圖1d),當長時間處理細胞時這一問題尤為突出,持續增殖、增殖停止或細胞死亡的綜合情況可能會發生。此外,只知道活細胞數或死細胞數難以了解藥物效果,例如,一個樣本起初有100個活細胞,后來有100個死細胞和0個活細胞,這與一個樣本起初有100個活細胞,后來有100個死細胞和400個活細胞是不同的。我們需要知道在實驗開始時的活細胞數以及在實驗結束時的活細胞數和死細胞數,才能清楚地了解特定治療的效果[3,4]。
Tip3:考慮時間對結果的影響
細胞死亡是隨時間推移而發生的過程,在最終導致細胞死亡信號到達和實際質膜破裂之間通常會發生一段信號轉導期,這個時間可以從幾分鐘、幾小時、或到幾天不等,這是致命刺激的性質[5](圖1d)影響而造成的。因此,重要的是要注意何時進行細胞死亡測量。通常會使用一個任意的終點(例如,6、24或48小時)。重要的是要考慮終點的選擇是否有生物學依據,以及如果提前或推遲測量結果會是怎樣。某些細胞死亡抑制劑或基因操作可能并不能完全阻止細胞死亡,而只是改變了細胞死亡的時間[3]。因此,理想情況下細胞死亡是隨著時間的推移而測量的。在采用終點檢測法時,合理選擇時間點至關重要。
在質膜破裂后的某個時刻,原始細胞物質可能會完全解體(圖1a),這可能將導致對細胞死亡的誤解。相對于細胞群中細胞死亡的開始時間而言,過晚地測量可能意味著只有少量死亡細胞能保持足夠的完整性,無法通過任何方式對其進行計數。如果一種擾動迅速殺死了一些細胞,但剩余的活細胞繼續增殖直到較晚的實驗終點,這可能會掩蓋早期的細胞死亡效應,并導致一種“幸存者偏差”,即特定處理的致死效應被忽視。對群體中的細胞死亡進行延時測量有助于改善這些問題[3,4]。
Tip4:考慮細胞增殖對數據解釋的影響(第一部分)
特別是在使用終點法測定細胞活力時,細胞增殖可能會影響對細胞死亡的檢測和解釋。[6]在對照條件下的活細胞數量通常會在實驗開始和結束之間會迅速增加。在已發表的研究中頻繁使用24或48小時作為時間終點的部分原因是需要在這些細胞本身密度過大導致細胞死亡之前從對照條件中捕獲數據(圖1d)。更重要的是,相對于任何處理過的細胞群體,對照細胞群體的無限增殖可能無意中導致被誤解為對細胞死亡的影響(圖1d)[7]。
細胞增殖不會因許多致命的刺激而停止,在細胞死亡開始前的幾個小時內,活細胞的數量反而會繼續增加(圖1d)[3]。另一方面,DNA損傷或生長因子信號的抑制可引起雙相反應,即細胞首先停止增殖,之后不再增殖的細胞發生死亡[8]。這些動態很難用任何類型的終點分析來捕捉且不能輕易區分增殖停止和細胞死亡的大量代謝。隨著時間的推移,直接計數活細胞和死細胞的技術可以幫助捕捉這些效應,并將細胞死亡歸一化到一個群體中觀察到的起始或max活細胞數[3,4]。還開發了一些特殊的指標來衡量細胞增殖和細胞死亡是如何協調的,并有助于解釋群體水平上的細胞死亡[7]。
Tip5:考慮細胞增殖對數據解釋的影響(第二部分)
同時檢查一種處理對許多不同細胞系的影響是非常常見的。細胞系之間細胞增殖速率的差異會很大程度地影響對這些數據的解釋。尤其在批量測定細胞活力時,相比于增殖較快的細胞,生長較慢的細胞通常對治療誘導的細胞死亡表現出更強的抵抗力。HER2陽性乳腺癌細胞往往比培養的其他乳腺癌細胞增殖得更慢,因此傳統的測量方法無法捕捉到這些細胞對HER2抑制劑的獨特敏感性。歸一化方法可以減輕生長速率變化的混雜效應,更準確地測量處理因素誘導的細胞死亡[8,9]。
Tip6:數據規范化
細胞活力或細胞死亡通常以標準化和簡化的模式呈現(例如0到1或0%到100%的細胞死亡)。當使用大量代謝物計算時,通常減去培養基或培養容器產生的背景信號來對結果進行歸一化處理,然后將實驗組與陰性對照組進行比對。“細胞活力”降低50%可能表示所有細胞的增殖速度減慢50%,也可能表示群體中某些細胞死亡而其他細胞存活并增殖。
在一些研究中,結果被進一步抽象為相對于對照組的“細胞死亡百分比變化”。如果沒有進一步的信息,就無法獲知細胞群中多少細胞死亡。“細胞死亡增加100%”可以解釋為2%和4%的死亡細胞之間的差異,也可能意味著40%和80%的死亡細胞之間的差異。這種形式的數據歸一化可能會影響我們對生物效應的判斷。其他的簡化方法,如以熱圖形式顯示匯總數據,可以簡明扼要地傳達許多觀測值。這些描述只顯示了平均值,不同條件下存在的生物變異性仍不清楚。有時,應用的歸一化和簡化程度越多,就越難直觀地理解一種處理方法如何影響細胞死亡。
Tip7:仔細檢查圖表標注
細胞死亡數據結果的呈現方式會影響我們對這些結果的解讀。細胞死亡分析通常以x-y圖的形式呈現,其中y軸是“細胞死亡”的某種測量值。衡量標準可能包括活細胞計數、死細胞計數、群體中檢測到的細胞死亡標記物的數量或群體代謝活動。需要特別注意這些不同類型的數據在y軸是如何標注的,尤其是來自不同檢測方法的數據被歸一化并轉換成“細胞死亡百分比”或“細胞存活百分比”值時。如上所述,大量代謝物讀數并不直接體現細胞死亡情況,但有時會被作為細胞死亡的衡量標準。同樣,僅對細胞死亡或死細胞情況進行定量也不一定能有效說明群體中細胞死亡的總體水平。理想情況下,y軸應標注實際的測量值,以避免數據被過度解讀或誤讀的風險。此外,y軸的范圍也很重要,有時y軸不是呈現一個完整的范圍(0-100%),而是被限制在一組有限的值之間,這往往會在視覺上放大差異。
Tip8:注意細胞菌株和細胞培養條件
細胞供應源、傳代數和生長條件是細胞死亡實驗中的重要變量。培養中的細胞在不斷變化,這種變化可能導致不同實驗室使用的同一細胞系菌株之間細胞死亡的差異[9]。細胞死亡反應也會因生長培養基成分、細胞數量或培養容器中的細胞匯合度不同而存在差異。因此,關注上述變量至關重要。在某些情況下,實驗之間、實驗室之間的結果差異可以用細胞系克隆漂移、檢測細胞融合差異及細胞生長條件來解釋[11,12]。雖然細胞死亡數據的報告程序在不斷改進,但是存在的變量以及差異并不是一直出現在所公布的方法內。
Tip 9:關注效應量和可重復性
可重復性是生物學研究中的重大難題。即使是相對簡單的實驗設計,也包含數十個潛在的誤差來源,這些誤差來源與原材料、試劑、細胞培養和接種條件、移液器校準等有關[13]。即使盡可能使用相同的試劑和實驗方案,不同實驗室的細胞存活率實驗結果仍然會有差異[11]。 重要的是要考慮到,在細胞死亡研究中,實驗誤差或隨機誤差可能是造成不同條件下觀察到差異的原因。
結果可重復性是避免誤差的一道重要防線。在細胞生物學和動物研究中,實驗可重復性究竟是什么呢?已發表的研究報告在這一點上并不明確。有些研究認為在多孔板的平行孔中同時進行的實驗是“獨立”的重復,這種方法節省了時間,但很大限度地降低了生物變異性和系統性產生的風險。有時實驗會在不同時間出結果,但只有一個獨立實驗的數據會作為代表性數據顯示出來,這就很難評估其生物變異程度。
細胞和試劑即使來自同一動物或同一批次,在不同時間進行重復實驗也會存在類似的潛在混淆風險。因此需謹慎對待規模較小、在獨立實驗中未重復且僅在一種環境中得到驗證的細胞死亡。在一種細胞系中獲得的細胞死亡發現是否也能在另一種細胞系中獲得,以及獨立和正交的方法(如細胞死亡機制特異性標記物的生化讀數和活細胞與死細胞成像計數)是否都指向相同的機制結論[14]也是關鍵問題之一。由兩個或更多個獨立研究小組得出的類似機制發現其可信度會更高,并且比僅在單個實驗室中獲得的任何結果更具有普遍性。
Tip 10:仔細評估細胞死亡機制
細胞死亡可以通過細胞凋亡或幾種非凋亡細胞死亡機制之一進行,這些機制并不容易區分。研究者有必要關注如何監測不同的細胞死亡機制。細胞死亡機制特異性標志物會報告一種細胞死亡機制的激活情況,但對可能同時發生的其他細胞死亡機制視而不見。我們漸漸深入發現了一些機制特異性細胞死亡標志物。例如,磷脂酰絲氨酸結合Annexin V歷來被用作細胞凋亡的標志物,但磷脂酰絲氨酸也可能暴露在經歷壞死性凋亡和其他形式的非凋亡細胞死亡的細胞表面[15]。在解釋使用機制特異性細胞死亡抑制劑獲得的結果時也需要謹慎,這可能會產生脫靶效應。例如一些小分子具有獨立于蛋白質結合的隱自由基捕獲抗氧化活性,可抑制鐵死亡[16]。通過不斷完善和更新用于評估細胞死亡機制的標準,有可能更清晰地解釋現有數據和新數據。
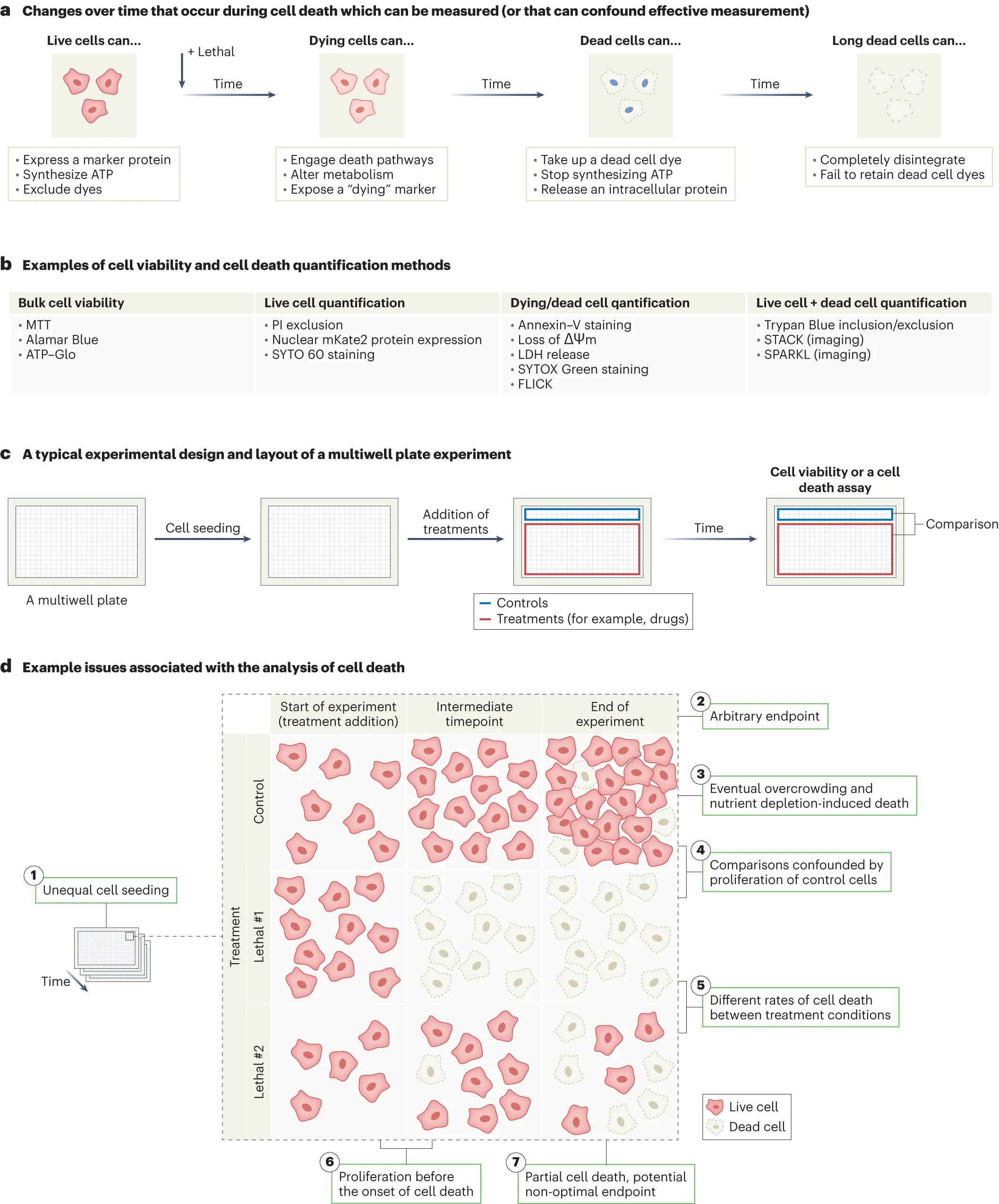
與細胞死亡檢測相關的問題概述
a、活細胞、瀕死細胞和死細胞可以產生獨特的和可測量的信號。b、常見細胞活力和細胞死亡定量方法舉例。FLICK,基于熒光和裂解的細胞死亡動力學推斷;LDH,乳酸脫氫酶;MTT,3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide;PI,碘化丙啶;SPARKL,使用實時動力學標記的單細胞和群體水平分析;STACK,可擴展的細胞死亡動力學延時分析。 c、多孔板中典型的細胞死亡檢測實驗設計示例。d、七種不同的變量和因素會影響細胞死亡實驗的解釋。在使用終點測量法和間接生化檢測法(如 ATP 總豐度或四氮唑還原強度)時,這些影響混淆數據解釋的可能性尤為嚴重。
那么有什么樣的工具可以有助于細胞死亡檢測呢?
奎克泰生物的EVE™和ADAM™系列自動細胞計數儀,操作簡單,快速區分活死細胞,計算細胞存活率!
奎克泰生物的JuLI™系列實時活細胞成像分析儀能夠放置于培養箱內,對細胞進行實時觀察、拍攝記錄生長周期的全過程,在保證細胞生長環境穩定的情況下,同時對細胞計數、拍照,形成視頻,分析融合度與生長曲線,提供細胞死亡量化結果!

[1]Dixon SJ, Lee MJ. Quick tips for interpreting cell death experiments. Nat Cell Biol. 2023 Dec;25(12):1720-1723.
[2] Chan GK, Kleinheinz TL, Peterson D, Moffat JG. A simple high-content cell cycle assay reveals frequent discrepancies between cell number and ATP and MTS proliferation assays. PLoS One. 2013 May 17;8(5):e63583.
[3] Forcina GC, Conlon M, Wells A, Cao JY, Dixon SJ. Systematic Quantification of Population Cell Death Kinetics in Mammalian Cells. Cell Syst. 2017 Jun 28;4(6):600-610.e6.
[4] Gelles JD, Mohammed JN, Santos LC, Legarda D, Ting AT, Chipuk JE. Single-Cell and Population-Level Analyses Using Real-Time Kinetic Labeling Couples Proliferation and Cell Death Mechanisms. Dev Cell. 2019 Oct 21;51(2):277-291.e4.
[5] Richards R, Schwartz HR, Honeywell ME, Stewart MS, Cruz-Gordillo P, Joyce AJ, Landry BD, Lee MJ. Drug antagonism and single-agent dominance result from differences in death kinetics. Nat Chem Biol. 2020 Jul;16(7):791-800.
[6] Harris LA, Frick PL, Garbett SP, Hardeman KN, Paudel BB, Lopez CF, Quaranta V, Tyson DR. An unbiased metric of antiproliferative drug effect in vitro. Nat Methods. 2016 Jun;13(6):497-500.
[7] Schwartz HR, Richards R, Fontana RE, Joyce AJ, Honeywell ME, Lee MJ. Drug GRADE: An Integrated Analysis of Population Growth and Cell Death Reveals Drug-Specific and Cancer Subtype-Specific Response Profiles. Cell Rep. 2020 Jun 23;31(12):107800.
[8] Hafner M, Mills CE, Subramanian K, Chen C, Chung M, Boswell SA, Everley RA, Liu C, Walmsley CS, Juric D, Sorger PK. Multiomics Profiling Establishes the Polypharmacology of FDA-Approved CDK4/6 Inhibitors and the Potential for Differential Clinical Activity. Cell Chem Biol. 2019 Aug 15;26(8):1067-1080.e8.
[9] Hafner M, Niepel M, Chung M, Sorger PK. Growth rate inhibition metrics correct for confounders in measuring sensitivity to cancer drugs. Nat Methods. 2016 Jun;13(6):521-7.
[10] Ben-David U, Siranosian B, Ha G, Tang H, Oren Y, Hinohara K, Strathdee CA, Dempster J, Lyons NJ, Burns R, Nag A, Kugener G, Cimini B, Tsvetkov P, Maruvka YE, O'Rourke R, Garrity A, Tubelli AA, Bandopadhayay P, Tsherniak A, Vazquez F, Wong B, Birger C, Ghandi M, Thorner AR, Bittker JA, Meyerson M, Getz G, Beroukhim R, Golub TR. Genetic and transcriptional evolution alters cancer cell line drug response. Nature. 2018 Aug;560(7718):325-330.
[11] Elliott JT, Rösslein M, Song NW, Toman B, Kinsner-Ovaskainen A, Maniratanachote R, Salit ML, Petersen EJ, Sequeira F, Romsos EL, Kim SJ, Lee J, von Moos NR, Rossi F, Hirsch C, Krug HF, Suchaoin W, Wick P. Toward achieving harmonization in a nano-cytotoxicity assay measurement through an interlaboratory comparison study. ALTEX. 2017;34(2):201-218.
[12] Haverty PM, Lin E, Tan J, Yu Y, Lam B, Lianoglou S, Neve RM, Martin S, Settleman J, Yauch RL, Bourgon R. Reproducible pharmacogenomic profiling of cancer cell line panels. Nature. 2016 May 19;533(7603):333-7.
[13] Rösslein M, Elliott JT, Salit M, Petersen EJ, Hirsch C, Krug HF, Wick P. Use of Cause-and-Effect Analysis to Design a High-Quality Nanocytotoxicology Assay. Chem Res Toxicol. 2015 Jan 20;28(1):21-30.
[14] Vaux DL. Research methods: Know when your numbers are significant. Nature. 2012 Dec 13;492(7428):180-1.
[15] Gong YN, Guy C, Olauson H, Becker JU, Yang M, Fitzgerald P, Linkermann A, Green DR. ESCRT-III Acts Downstream of MLKL to Regulate Necroptotic Cell Death and Its Consequences. Cell. 2017 Apr 6;169(2):286-300.e16.
[16] Shah R, Shchepinov MS, Pratt DA. Resolving the Role of Lipoxygenases in the Initiation and Execution of Ferroptosis. ACS Cent Sci. 2018 Mar 28;4(3):387-396.



