
文獻解讀:相關多尺度冷凍成像揭示新冠病毒的組裝和排出

內容摘要
自SARS-CoV-2大流行爆發(fā)以來,對純化的病毒成分和滅活病毒進行了深入的結構研究。然而,關于SARS-CoV-2感染如何在天然細胞環(huán)境中進行的結構和超微結構證據很少,并且缺乏關于SARS-CoV-2復制周期的全面知識。為了將SARS-CoV-2誘導的細胞病變事件與冷凍水合細胞中的病毒復制過程相關聯(lián),我們建立了一個獨特的多模式、多尺度冷凍相關平臺,以對Vero細胞中SARS-CoV-2感染進行成像。該平臺將連續(xù)低溫FIB/SEM體積成像和軟X射線低溫層析成像與基于細胞薄片的低溫電子層析成像(CryoET)和亞同步圖平均相結合。在這里,我們報告了關鍵的SARS-CoV-2結構事件,例如病毒RNA轉運入口、病毒組裝中間體、病毒出口途徑和天然病毒刺突結構,在整個細胞體積的背景下,揭示了劇烈的細胞化學變化。這種綜合方法允許從整個細胞到單個分子全面觀察SARS-CoV-2感染。
自2019年12月以來,世界一直處于被稱為“本世紀最大的流行病”的中期。病原體是嚴重急性呼吸綜合征冠狀病毒2(SARS-CoV-2),由此引發(fā)的疾病是2019年冠狀病毒病。冠狀病毒是具有陽性非分段RNA基因組的小型包膜病毒。在RNA病毒中,冠狀病毒擁有最大的基因組之一,它們的復制是復雜的,包括移碼和復制酶跳躍,產生大量的RNA雙鏈體。冠狀病毒和大多數RNA病毒一樣,誘導一系列膜室的發(fā)育,這些膜室隔離并保護病毒成分,從而提高復制效率和先天免疫識別逃逸。
所有冠狀病毒結構蛋白都是由正義亞基因組RNA片段的翻譯產生的,正義亞基因RNA片段是由負義亞基因組RNAs片段產生的,而負義亞基因RNAs片段又是由病毒基因組轉錄時的復制酶跳躍產生的。其中,S蛋白構成病毒刺突,負責細胞附著、進入和融合。它采用兩種主要構象:融合前,由亞基S1和S2的三聚體組成;融合后,僅由S2組成的非活性構象。N蛋白負責包裹和保護基因組病毒RNA(vRNA),形成駐留在病毒顆粒內部空間的核糖核蛋白復合物。E蛋白是最小的結構蛋白,被認為是離子通道。最后,M蛋白在SARS-CoV-2中含量最高,是一種跨膜蛋白,分布在病毒脂質膜的內表面。
在這項研究中,我們利用了一種獨特的相關多模態(tài)多尺度冷凍成像方法來研究在接近自然條件下Vero細胞中SARS-CoV-2的復制。該方法使SARS-CoV-2感染從整個細胞到單個病毒刺突分子的整體觀點成為可能,揭示了SARS-CoV-2組裝的途徑以及SARS-CoV-1感染的出口和細胞病變效應。這些結果證實了之前通過對染色的塑料包埋樣品進行薄片電子顯微鏡(EM)和連續(xù)聚焦離子束掃描電子顯微鏡(FIB/SEM)獲得的發(fā)現(xiàn),并進一步擴展了我們對SARS-CoV-2組裝和出口以及其在其他膜室中的存在的了解。它驗證了連續(xù)低溫FIB/SEM和軟X射線斷層掃描作為研究與高分辨率低溫電子斷層掃描(cryoET)相關的全細胞形態(tài)的技術,具有冷凍水合條件和快速直接制備樣品的優(yōu)勢。
實驗方法
細胞系和病毒
將非洲綠猴腎Vero Cc-81細胞(雌性)(ATCC,Ccl-81)保存在補充有5%胎牛血清、10單位/mL青霉素(Gibco)、10µg/mL鏈霉素(Gibco)和2mM L-谷氨酰胺的默克高糖培養(yǎng)基(D-MEM)中。細胞系未經驗證。SARS-CoV-2分離株Beta CoV/England/02/2020(EPI_ISL_407073)由Maria Zambon教授貯藏,并通過BEI資源、NIAID、NIH:SARS相關冠狀病毒2,分離株England/02/2020NR-52359獲得。本研究中使用的病毒原液已通過Vero Ccl-81細胞中的菌斑測定進行滴定,并進行測序,以確認通過擴增傳代保留了核苷酸呋喃裂解位點(FCS)。
樣品制備
Vero Ccl-81細胞(ATCC)保持如上所述。將16000個細胞接種在六孔板孔中經纖維連接蛋白處理的G300F1 R2/2金電鏡載網的碳膜側。使用SARS-CoV-2 England/02/2020的第3代進行感染,MOI為0.5、0.1或0(陰性對照)。從Vero Ccl-81細胞(ATCC)中取出培養(yǎng)基,并用1%胎牛血清、10單位/mL青霉素(Gibco)、10µg/mL鏈霉素(Gibco)和2 mM L-谷氨酰胺(Gibco)稀釋在0.5mL D-MEM高糖培養(yǎng)基(Merck)中的適量病毒替換。將細胞在室溫下孵育15分鐘,然后向每個孔中再加入1.5mL培養(yǎng)基。然后將平板在37°C下孵育24小時,然后丟棄上清液并用2 mL PBS洗滌細胞。然后通過在PBS中加入3mL 4%多聚甲醛在室溫下固定細胞1小時。固定后,將載網放置在徠卡GP2中,將1µl BSA(EMS)中的濃縮10nm金基準點施加到電鏡載網的背面,并從背面進行印跡,印跡后將載網浸入液態(tài)乙烷中冷凍。玻璃化載網儲存在液氮中,直到收集數據。
CryoET數據采集
傾斜系列采集是在FEI Titan Krios G2(Thermo Fisher Scientific)透射電鏡上進行的,該電鏡可在300kV下操作,并配備了能量過濾器和post-GIF K3探測器。使用SerialEM 3.8版傾斜系列控制器記錄傾斜系列,完整細胞的像素大小為1.63、2.13和4.58Å,薄片的像素大小為2.13和7.58Å。零損耗成像用于具有20eV狹縫寬度的所有傾斜系列。散焦值范圍為−2至−7µm,但使用50µm散焦的7.58Å像素大小的薄片除外。插入100µm物鏡光闌。所有傾斜系列均采用分組劑量對稱方案;在+/-60度范圍內以3度增量收集完整細胞,每組3個,總劑量為120–135 e/Å2;對于薄片,在+/-54度范圍內以3度增量收集,在4.58Å時為每組3個,總劑量為120–135 e/Å2,在7.58 Å時為每組10個,總劑量為70–90 e/Å2。在每次傾斜時進行自動聚焦和跟蹤,并以10Å/s的目標速率在傾斜反轉時進行漂移測量。在每次傾斜時,在超分辨率模式下使用相關雙采樣記錄5個影片幀,并以lzw壓縮tif格式保存,無增益歸一化。影片隨后在運動校正期間進行增益歸一化,并將傅里葉裁剪回物理像素大小。在每個傾斜系列之后,運行腳本以獲取新的暗參考并重置散焦偏移。總共收集了294個傾斜序列,其中56個傾斜序列來自未感染的對照細胞(20個來自細胞薄片,36個來自細胞外周),238個傾斜序列源自SARS-CoV-2感染的細胞(90個來自薄片,148個來自細胞外周)。
CryoFIB薄片制備
SARS-Cov-2感染細胞的薄片銑切是使用配備有PP3010冷凍傳輸系統(tǒng)(Quorum Technologies)的Scios DualBeam CryoFIB(ThermoFisher Scientific)和使用xT v7.6軟件(ThermoFisher Scientific)進行的。在PP3010制備腔室內保持−175°C冷臺上對載網進行濺射鍍膜。在傳輸轉載到Scios腔室內−168°C冷臺后,使用SEM(在5kV和13pA下操作)檢查載網,并鑒定出與透射電子顯微鏡(TEM)相關的感染細胞。使用氣體注入系統(tǒng)[三甲基(甲基環(huán)戊二烯基)鉑(IV),ThermoFisher Scientific]涂覆載網表面3秒,形成厚度約為3µm的鉑保護層。使用在30kV下操作的離子束和從300到30pA遞減的電流進行銑切。在30pA下,薄片厚度小于300nm。在銑切的最后階段,在2kV和13pA下對薄片進行SEM檢查。
系列CryoFIB/SEM體積成像
樣品在配備有Quorum PP3010冷凍制備傳輸系統(tǒng)的Zeiss Crossbeam 550XL雙束電鏡上成像。將它們裝在定制樣品架上,并在Quoum冷凍制備腔室的冷臺上以10mA電流濺射鍍鉑60秒,然后傳輸轉移至電鏡冷臺。電鏡冷臺溫度設置為−165°C,同時防污染冷阱保持在−185°C。在Pt氣體儲存保持在25°C的前提下,使用FIB-SEM的內部GIS系統(tǒng),再次用Pt涂覆2×30 s后,然后以45°傾斜對樣品進行成像。在30 kV、7 nA、15μm寬度條件下銑切出初始梯形溝槽,最終深度為10μm,以30 kV和1.5 nA條件對深度為10μm的矩形面進行精銑。連續(xù)切片和圖像采集如下:使用30 kV、700 pA探針電流設置FIB銑切,z切片步進為20 nm,貫穿深度為10µm的整個研磨面;SEM成像是在3024×2304像素數量下進行,這導致像素尺寸為6.5nm,電子束設置為2kV,35pA,束駐留時間100ns,掃描速度1,將100行掃描上的信號平均化作為降噪策略。使用的軟件版本為Zeiss SmartSEM v6.06和Zeiss SmartFIB v1.11.10。

CryoET image processing,Subtomogram averaging,
Soft X-ray cryo-tomography,Quantification and statistical analyses and so on
(限于篇幅從略,僅呈現(xiàn)數據圖表)
結論
SARS-CoV-2復制在宿主細胞中誘導深刻的細胞病變效應。為了在接近生理條件下對細胞內體系中SARS-CoV-2的復制進行成像和研究,我們用SARS-CoV-2以0.1和0.5倍感染率(MOI)去感染在編碼的電鏡載網上生長的Vero細胞。在感染后24小時(hpi),將細胞用4%多聚甲醛固定并浸入液體乙烷中冷凍。如工作流程所示(補充圖1),首先在Titan Krios中對含有SARS-CoV-2感染細胞的電鏡載網進行成像,以通過檢測細胞外圍的大量病毒顆粒來識別感染細胞(MOI為0.1時為39.2%,MOI為0.5時為45.4%)。Vero細胞中SARS-CoV-2復制動力學小于24小時。因此,我們的樣本可能包含不同復制周期階段的感染細胞混合物。在感染細胞的外圍收集CryoET傾斜系列。然后將載網轉移到FIB/SEM雙束電鏡上,用連續(xù)冷凍FIB/SEM體積成像法對相同的感染細胞進行成像,或冷凍FIB銑切目標區(qū)域的細胞薄片,以便收集額外的cryoET傾斜系列。同時,我們通過軟X射線冷凍斷層掃描對其他感染細胞進行了成像。受感染和未受感染細胞的定量和相關數據集,以及不同觀察結果的定量分析,在補充表1中予以總結。這些成像模式在不同長度尺度上提供了必要的結構和超微結構信息,以便在其細胞環(huán)境中可視化感染病毒,并且具有高度互補性。事實上,這種獨特的方法能夠以多模式、多尺度和相關的方式直接可視化SARS-CoV-2復制和細胞病變效應。
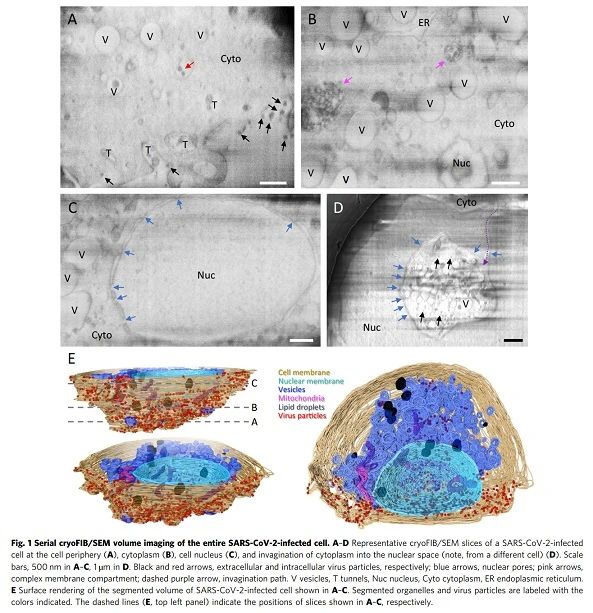
與未感染細胞相比(補充圖2和補充影片1),五個SARS-CoV-2感染細胞的連續(xù)冷凍FIB/SEM全細胞成像顯示了廣泛的細胞病理學改變,如圖1和補充影片2-6所示。在細胞外圍,有許多電子致密顆粒,其大小與SARS-CoV-2病毒一致(圖1A,黑色箭頭)。觀察到許多膜隧道狀結構從細胞膜向細胞質深入細胞,含有病毒樣顆粒(圖1A,“T”和補充影片2)。這些隧道狀結構類似于在HIV-1感染的巨噬細胞中觀察到的結構,病毒通過這些結構離開感染的細胞。此外,在與細胞膜不相連的細胞內小泡中也發(fā)現(xiàn)了電子密集的病毒樣顆粒(圖1A,紅色箭頭)。在細胞深處,我們發(fā)現(xiàn)大部分細胞質(圖1B),尤其是核周區(qū)域(圖1C)被大量小泡“V”占據(補充影片2-5)。SARS-CoV-2感染細胞和未感染細胞中的核孔清晰可辨(圖1C和補充圖2A,C,藍色箭頭)。我們在感染的細胞中觀察到電子致密的復合膜室(圖1B,粉紅色箭頭)。先前已經觀察到類似的結構,并將其描述為在染色的塑料包埋細胞的薄切片中的大型含病毒囊泡(LVCV)。與對照細胞相比,在感染細胞中觀察到的一個更顯著的特征是細胞核的細胞病變損傷,在極端情況下,近一半的細胞核被內陷的細胞質占據(圖1D,補充影片6和補充圖3G)。在SARS-CoV-2感染的肺細胞和腸細胞的染色塑料切片的電鏡觀察中也注意到這種細胞質內陷,這表明SARS-CoV-2感染具有強烈的細胞病變效應。
利用軟X射線冷凍斷層掃描提供的高通量全細胞成像能力,我們分析了5個感染細胞,這些細胞通過冷凍ET成像在細胞外圍識別并確認SARS-CoV-2感染,另外還有12個來自對照網格的未感染細胞。在整個細胞水平上,軟X射線圖像顯示了線粒體的實質性變化,因為在未感染細胞的整個細胞質中存在長管狀線粒體(補充圖3A,C,黃色箭頭),在感染細胞中很少見到(補充圖3B,D)。我們觀察到SARS-CoV-2感染細胞中線粒體的數量顯著減少(SARS-CoV感染細胞中每斷層圖像15.4±10.2,對照細胞中91.3±32,p=0.0001;補充圖4A)。這與先前描述SARS-CoV-2感染肺細胞導致線粒體網絡改變的研究結果一致。對三個感染細胞和兩個未感染細胞的人工分割的全細胞冷凍FIB/SEM體積的測量表明,SARS-CoV-2感染細胞中線粒體的長度明顯短于對照細胞(SARS-CoV-2感染細胞的線粒體長度為0.60±0.70μm,對照細胞為0.90±1.14μm;p=0.0012;補充圖4B),線粒體體積沒有顯著減少(SARS-CoV-2感染的細胞為0.058±0.196μm3,對照細胞為0.141±0.763μm3,p=0.113;補充圖4C),表明線粒體形態(tài)發(fā)生了變化。我們還在核周區(qū)域觀察到許多小泡(補充圖3F),其大小與感染細胞中的DMV和細胞質內陷(補充圖3G)一致。總之,這些冷凍成像結果證實并擴展了先前與SARS-CoV-2感染相關的細胞病理學改變的發(fā)現(xiàn)。
討論
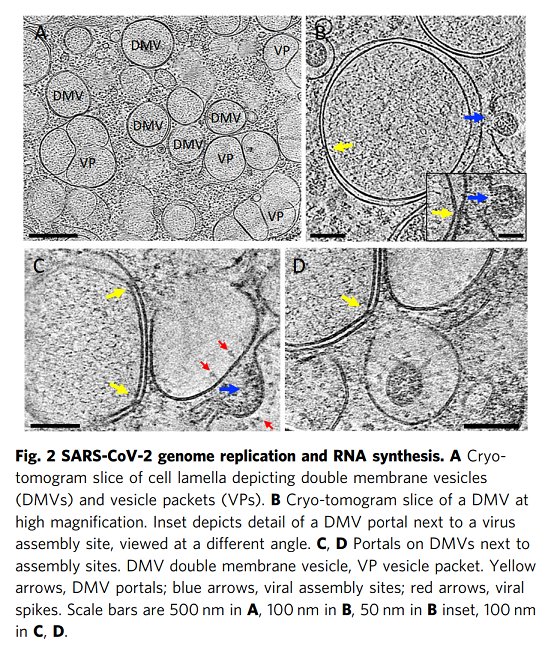
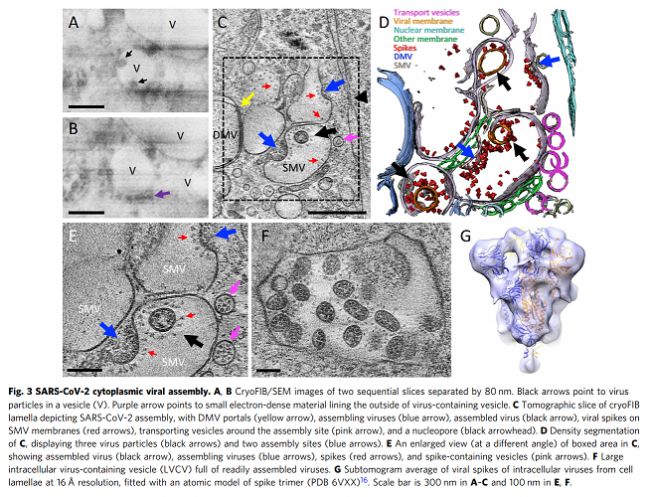
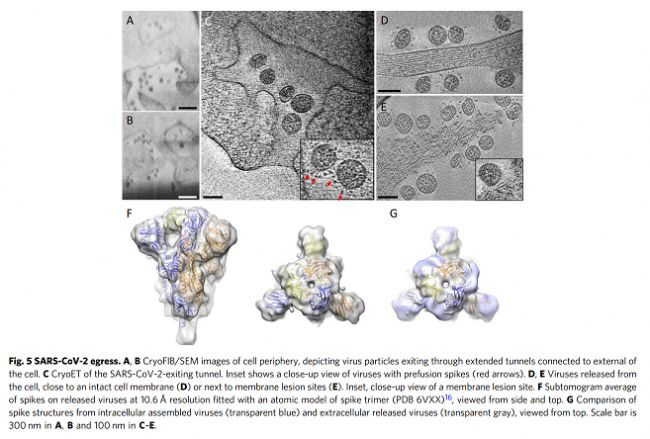
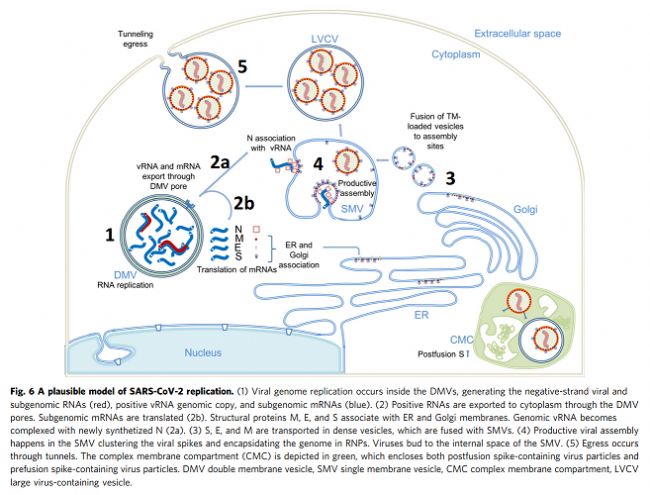
我們使用相關方法對近天然玻璃化細胞中SARS-CoV-2感染進行成像,包括從整個細胞水平到亞細胞和分子水平的多個空間尺度。這種方法產生了感染過程的整體視圖,描述了病毒誘導的細胞病變及其與原位結構的相關性。至關重要的是,通過該工作流實現(xiàn)的多尺度成像數據的整合(補充圖1)使我們能夠在感染細胞的細胞病變后果的背景下直接可視化SARS-CoV-2復制的結構事件。鑒于先前研究中對SARS-CoV-2和其他冠狀病毒的先驗知識,這些圖像使我們提出了SARS-CoV-2復制的增強模型,特別是病毒基因組復制、組裝和出口,如下所述。SARS-CoV-2復制過程的圖像顯示,它是一個空間組織良好且高效的過程。從基因組復制到蛋白質合成和運輸,再到病毒組裝和出芽,每一步都發(fā)生在特定目的的緊密聯(lián)系的細胞質隔間中。如圖6所示,RNA復制,包括基因組vRNA和亞基因組mRNA,發(fā)生在DMV中,將其從宿主細胞固有免疫應答中分離出來(步驟1)。然后,新合成的vRNA通過跨膜入口從DMV運輸到這些DMV附近的病毒組裝位點(步驟2a),而mRNA通過相同的入口排出到細胞質,隨后轉移到ER/Golgi以產生蛋白質(步驟2b)。在ER/Golgi網絡中產生的三聚體預融合形式的病毒尖峰通過小的運輸囊泡運輸到組裝位點(步驟3)。這些囊泡與SMV膜融合,其中在存在vRNA和N蛋白的情況下,病毒尖峰聚集在組裝位點,導致正的膜彎曲,并最終將病毒粒子萌發(fā)到SMV中(步驟4)。組裝和出芽過程產生含有病毒的囊泡,囊泡可以含有多種病毒顆粒。然后,病毒顆粒可以通過通道排出,可能通過溶酶體胞吐(步驟5)。該模型的某些方面仍然是推測性的,需要在未來的研究中進行進一步的實驗驗證。
對SARS-CoV-2病毒在細胞內傳播過程中的基因組復制、組裝和排出過程進行多尺度觀察,對于促進我們對這種病原體的理解至關重要,因為它具有闡明新信息和激發(fā)醫(yī)學干預以阻止生產性感染的手段。這一過程的許多方面有待進一步研究,以剖析SARS-CoV-2復制的分子機制,包括其他病毒蛋白(如M和E)以及宿主蛋白和核心機構的作用。這項研究提供了SARS-CoV-2復制周期在接近自然條件下的直接觀察,以及直接來自細胞組裝和細胞外釋放的病毒顆粒的融合前尖峰的結構。通過這項研究開發(fā)的獨特方法和工作流程可廣泛應用于SARS-CoV-2以外的其他人類病原體感染過程的研究。
科學家團隊簡介




