
FV3000ЦъБҰЦРҝЖФәДXЦЗЧҝФҪЦРРД¶ЕҫГБЦҲFк ҪТКҫН»У|°lУэөД•ғТ№№қВЙРФ

ЦРҮшҝЖҢWФәДXҝЖҢWЕcЦЗДЬјјРgЧҝФҪ„“РВЦРРД
ЙъОпзҠНЁЯ^…fХ{Я@Р©ЙъАнәНРРһйЯ^іМЈ¬К№„УОпДЬүтЯm‘ӘИХіЈӯhҫіөДЧғ»ҜЎЈө«КЗЈ¬іэБЛФЪЦTИз°ЯсRф~УЧф~өДјҡ°ыЦЬЖЪ[5,6]Ўў№ыПүөДУр»Ҝ[7]Ј¬ТФј°ІёИй„УОпДIЕKЖч№ЩРОіЙ[8]ЦРУРЙЩБҝҲуөАНвЈ¬ИЛӮғҢҰУЪЙъОпзҠКЗ·сТФј°ИзәОХ{№қЙъОпуwФзЖЪ°lУэЯ^іМөДХJЧR…T·ҰЎЈ

ұҫСРҫҝөДСРҫҝИЛҶTФЪЗ°ЖЪөД№ӨЧчЦРҪЁБўБЛМШ®җҳЛУӣТ•ҫWДӨ-Т•н”ЙwН»У|өДЮD»щТт°ЯсRф~ДЈРН[9]Ј¬ФЪұҫСРҫҝЦР„tКЗАыУГФ“ДЈРНҒнМҪҫҝН»У|°lЙъКЗ·сҫЯУР•ғТ№№қВЙРФЎЈ
КЧПИЈ¬СРҫҝИЛҶTҢўіцЙъәуөД°ЯсRф~УЧф~·ЕФЪХэіЈББ°ө№вХХ—lјюПВЈ¬ҪӣЯ^ЛДМмөДУ–ҫҡ»т ҝТэЈЁentrainmentЈ©әуЈ¬й_Х№ТФ6РЎ•rһй•rйgйgёфЎўіЦАmғЙМмөДйL•rіМіЙПсЎЈ·ЦОц°l¬FН»У|”өБҝөДФцйLЛЩВКҙжФЪ°ЧМмҝмЎўТ№НнВэөД•ғТ№ХсКҺЈЁҲD1a,bЈ©ЎЈЯ@Т»ХсКҺ¬FПуФЪББ°ө№вХХ ҝТэәуЮDИліЦАmББ»тіЦАm°өөД№вХХ—lјюЈЁҲD1cЈ©Ј¬»тХЯҪӣЯ^·ҙПтөД°өББ№вХХ—lјюЈЁҲD1dЈ© ҝТэәу¶јДЬұЈіЦЈ¬¶шФЪЙъОпзҠ»щТтН»ЧғуwЈЁclocka-/-Ј©ЦРПыК§ЈЁҲD1eЈ©Ј¬МбКҫЯ@Т»ХсКҺКЬөҪғИФҙЙъОпзҠПөҪyХ{№қЈ¬ҫЯУР•ғТ№№қВЙРФЎЈ
ЯMТ»ІҪЈ¬СРҫҝИЛҶTНЁЯ^ёЯ•rйg·ЦұжВКіЙПс·ЦОцН»У|°lУэөД„У‘BЯ^іМЈ¬°l¬FЯ@Т»№қВЙКЗУЙН»У|РОіЙЈЁsynapse formationЈ©¶ш·ЗН»У|ПыіэЈЁsynapse eliminationЈ©ЛЩВКөД•ғТ№Іо®җФміЙөДЈЁҲD1fЈ©ЎЈ
ҪУЦшЈ¬СРҫҝИЛҶT°l¬FН»У|ЙъйLЛЩВКөД№қВЙРФФЪПВЗрДXКіУыЛШДЬЈЁhypocretin/orexinЈ©ЙсҪӣПөҪyЈЁҲD1gЈ©№ҰДЬЧи”аәуПыК§ЈЁҲD1hЈ©Ј¬МбКҫФ“ПөҪy…ўЕcХ{№қН»У|°lУэ№қВЙ¬FПуЎЈЧоәуЈ¬СРҫҝИЛҶTНЁЯ^ЙъОпзҠ»щТтН»Чғ»тКіУыЛШДЬЙсҪӣФӘКЬуwН»ЧғЖЖүДН»У|°lЙъөД№қВЙРФЈ¬°l¬FҝЙТФФміЙТ•ҫWДӨЙсҪӣ№қЙсҪӣФӘЭSН»·ЦЦҰЧғҙуЎўҸНлs¶ИҪөөНЈ¬Н»У|”өБҝңpЙЩЎўФЪЭSН»·ЦЦҰ…^УтЦРөДГЬ¶ИҪөөНәНЕЕІјІ»ҫщәвөДҪYҳӢЧғ»ҜЈЁҲD1iЈ©Ј»Н¬•rНЁЯ^ҷzңyН»У|әуТ•н”ЙwЙсҪӣФӘөДёРКЬТ°Ј¬°l¬FЖдёРКЬТ°°lЙъБЛЕcН»У|З°ЙсҪӣФӘЭSН»·ЦЦҰЧғҙуПаәф‘ӘөДФцҙу¬FПуЈЁҲD1jЈ©Ј¬јҙЖдТ•дJ¶И№ҰДЬ°lУэКЬөҪУ°н‘ЎЈ

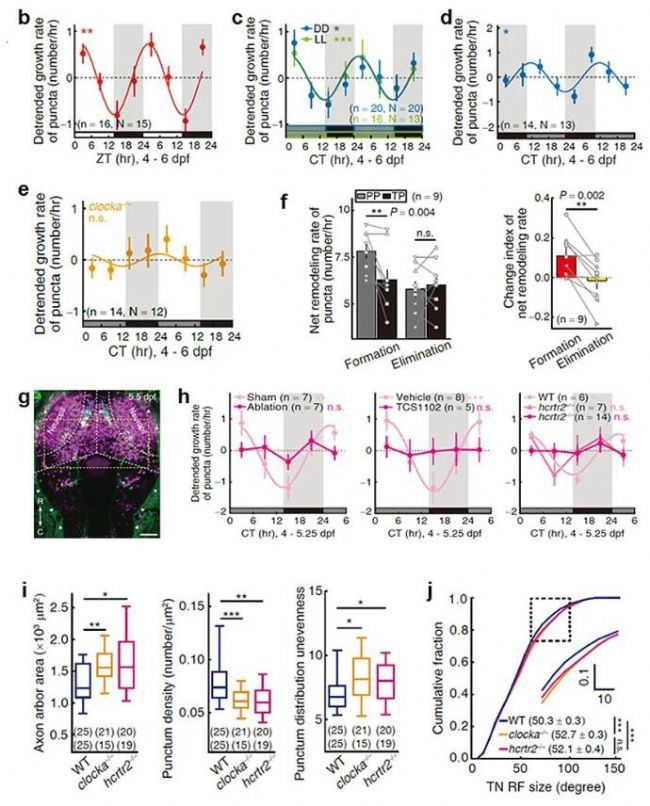

ҲD1 °lУэФзЖЪН»У|°lЙъөД•ғТ№№қВЙРФ№ӨЧчөДЦчТӘҪY№ыәН№ӨЧчДЈРН
ҙЛСРҫҝ№ӨЧчТ»·ҪГжҪТКҫБЛЙъОпзҠҢҰЙъОпуwФзЖЪ°lУэЯ^іМТІҫЯУРХ{№қЧчУГЈ¬”UХ№БЛЙъОпзҠөД№ҰДЬЧVЈ»БнТ»·ҪГжЈ¬ҪТКҫБЛЙсҪӣПөҪyӯhВ·°lУэЯ^іМЦРөДТ»·NРВөДХ{№қҷCЦЖЎЈ№ӨЧчЦР°l¬FөД…ўЕcЛҜГЯ-УXРСХ{№қөДПВЗрДXКіУыЛШДЬЙсҪӣФӘЧчһйЙъОпзҠөДПВУО…ўЕcХ{№қҙЛ№қВЙ¬FПуЈ¬һйСРҫҝЙъОпзҠЕcЛҜГЯ-УXРСөДПа»ҘЧчУГТФј°ЙъОпзҠөДЭ”іцВ·ҸҪМṩБЛРВөДСРҫҝЛјВ·ЈЁҲD1kЈ©ЎЈ

FV1000ХэЦГ

FV3000ХэЦГ
ФӯОДЈә
Circadian regulation of developmental synaptogenesis via the hypocretinergic system | Nature Communications
…ўҝјОД«I
1. Musiek, E. S. & Holtzman, D. M. Mechanisms linking circadian clocks, sleep, and neurodegeneration. Science (80-. ). 354, 1004–1008 (2016).
2. Thosar, S. S. & Shea, S. A. Circadian control of human cardiovascular function. Curr. Opin. Pharmacol. 57, 89–97 (2021).
3. Scheiermann, C., Gibbs, J., Ince, L. & Loudon, A. Clocking in to immunity. Nat. Rev. Immunol. 18, 423–437 (2018).
4. Reinke, H. & Asher, G. Crosstalk between metabolism and circadian clocks. Nat. Rev. Mol. Cell Biol. 20, 227–241 (2019).
5. Dekens, M. P. et al. Light regulates the cell cycle in zebrafish. Curr Biol 13, 2051–2057 (2003).
6. Dickmeis, T. et al. Glucocorticoids play a key role in circadian cell cycle rhythms. PLoS Biol. 5, 854–864 (2007).
7. Myers, E. M., Yu, J. & Sehgal, A. Circadian Control of Eclosion: Interaction between a Central and Peripheral Clock in Drosophila melanogaster. Curr. Biol. 13, 526–533 (2003).
8. Dan, H., Ruan, T. & Sampogna, R. V. Circadian Clock Regulation of Developmental Time in the Kidney. Cell Rep. 31, 107661 (2020).
ҫҙХҲьc“фҙЛМҺІйҝҙФӯОДЎЈ
В“ПөлҠФ’Јә4009690456
E-mailЈәmarketing.cn@evidentscientific.com

- ҠWӮҘөЗDSX2000ЕcOLS5100п@ОўзRЦъБҰ№влҠЮD“QөДЩ|БҝұOҝШЕcР§ДЬғһ»Ҝ
- К№УГEVIDENT№ІҫЫҪ№п@ОўзRҢҰСӘ№Ь»ҜД[Бцјҡ°ыЗтРҫЖ¬ЯMРРУ^ІмСРҫҝ
- EVIDENTп@ОўПөҪyИ«ЗтёчҙуЖЪҝҜҪьЖЪ°lұнөДХ“ОД…RҝӮ
- ҪьјtНвјӨ№вЖчЕcSilVIRМҪңyЖчЦъБҰёЯјү¶аЙ«ҹЙ№в№ІҫЫҪ№іЙПс
- EVIDENT№ІҫЫҪ№п@ОўзRЦъБҰЦРҮшҹбҝЖФәҹбҺ§ЧчОпСРҫҝ
- п@ОўзRРФДЬұOңyЖчЦъБҰіЙПсЦРРДЖҪЕ_ҙ_ұЈ¶ЁБҝҹЙ№віЙПсөДңКҙ_РФ
- і¬ёЯ·Цұжп@ОўзRөД°lХ№ј°ЖдФЪЙъОпбtҢWоIУтөД‘ӘУГ
- °ў –ҙДәЈД¬ІЎЙсҪӣГвТЯПа»ҘЧчУГөДҝХйg·ЦОц
- ЦұІҘЈә№ІҫЫҪ№ҹЙ№вүЫГьіЙПсФЪЦІОпПакPСРҫҝЦРөД‘ӘУГ
- EVIDENTҶўУГЦРОДЖ·ЕЖ"ҠWӮҘөЗ"Ј¬јУЛЩЦРҮшЯ\ IұҫНБ»Ҝ
- EVIDENTҹЁРВББПајҡ°ыДк•юТФ"ҠWӮҘөЗ"Ц®ГыјУЛЩұҫНБ»Ҝ
- ІМЛҫХэКҪ°lІјLightfield 4DёЯЛЩуwіЙПсјјРg
- EVIDENT 2024ДкҪKұPьcЈ¬ЙъГьҝЖҢWәН№ӨҳIЕ_ҡvСыДъДГ
- ЦұІҘоAёжЈәҹЙ№вүЫГьп@ОўзRіЙПсФӯАнј°‘ӘУГЯMХ№
- ЙӯОчЩҗЦЗҝЖјј2024ДҪДбәЪЙПәЈ·ЦОцЙъ»ҜХ№ҫ«ІК»Шоҷ
- EVIDENTөЪОеҢГИ«Зтп@ОўҲDПсҙуЩҗ«@Ә„ЧчЖ·ҪТ•Ф


