
PROTACˇÖ×ĶÔOĶÖŽ°Đĩ°°×ÅäķwĩÄßxņˇŊˇ¨
Į°ÎÄÎŌŌŅŊŊéŊBß^ŖŦPROTAC ˇÖ×ĶĶÉ°Đĩ°°×ÅäķwĄĸE3 ßBŊĶøÅäķwēÍÉÕßÖĐégĩÄßBŊĶļÎŊMŗÉŖŦÔOĶ PROTAC ÖŽŗõŖŦČũÕßĒÔˇÖeŋŧÁŋĄŖ
Ąö °Đĩ°°×Åäķw
ÔÚëHÔOĶš¤×÷ÖĐŖŦÍųÍųĪČūßxņēÃÄŋËĩ°°×ĄŖ°Đĩ°°×ĩÄĖôßxŋÉŌÔÄŌÔĪÂ×ücŋŧ]Ŗē
1ĄĸÔĩ°°×ĘĮˇņžßĶĐĖØŽĐÔ, ˇĮĖØŽĐÔĩ°°×ūÔėŗÉðĐļžĐÔŖģ
2ĄĸÔĩ°°×ĘĮˇņŲĶÚ PROTAC ˇÖ×ĶŗŖŌĩÄŊĩŊâĩ°°× (PROTACable °ĐücÔöŧĶ PROTAC ÔOĶĩÄĶĐЧĐÔ), 2021 ÄęŖŦNature Reviews Drug Discovery ësÖžķ§ÁËŌģÆĒĀûĶà PROTACtability ˇŊˇ¨Ôurĩ°°×°ĐücĩÄŋÉPROTACableĩÄÎÄÕÂŖŦÎÄÕÂ×îēķĖáŗöÖĩĩà PROTAC ģ¯ĩÄ°Đüc°üĀ¨ÁËŧ¤Ã¸ (MEKĄĸKRASĄĸCDK ēÍ Bcr/Abl)ĄĸŪDäŌō×Ķ (Čį p53ĄĸSTATĄĸRARĄĸER ēÍ AR)ĄĸąíĶ^ßz÷Ōō×Ķ (Čį HDAC ēÍ BET äåŊYĶō)ēÍ E3 ßBŊĶøąžÉí (Čį MDM2) ĩČĄŖ
3ĄĸÔĩ°°×ĘĮˇņĶĐĪāęPĩÄÅäķwģōž§ĐÍķĩĀŖŦß@ĘĮŋŧ]ĩŊÉĖIģ¯ÔOĶ PROTAC ĩÄЧÂĘĐÔŖŦČįÄŋĮ°ÅR´˛ļūÆÚĩÄÉŌŅÖĒŊYĩÄ ARV-471ēÍ ARV-766 ŊYˇÖeÔ´×ÔĶÚ Tamoxifen ēÍ (+)-JQ-1ĄŖ

ÉĪĘöČũücžųM×ãrŖŦŗŖŌĩÄ PROTAC ÔOĶąãŋÉßMĐĐŖēÄ°ĐücŌŅÖĒĩÄĪāęPÅäķwˇÖ×ĶЧęPĪĩŋŊYŗöËü˛ģĶ°íŊYēĪĩÄßmēĪ¸ÄÔėĩÄÎģÖÃŖŦģōĘĮĶĐĩ°°×ĩÄž§ķwÄŖĐÍŋÉąģĶÃĶÚĖMēYßxÄļøŋsĐĄēķĀm¸ßͨÁŋēYßxĩġļúĄŖ
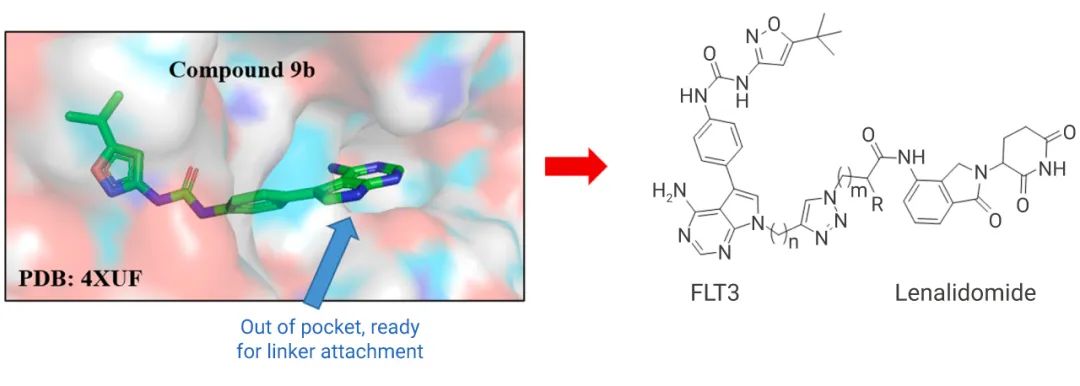
ŽČģŖŦÔÚëHÔOĶÖĐŖŦÍųÍųßūĶöĩŊ°Đĩ°°×ąžÉíĩÄČÔ´ĐÔÅäķwŖēļāëÄĄĸDNAĄĸĖĮæĩČŖŦģųĶÚ´ËŌ˛ĶĐĪāęPĩÄ p-PROTAC ąģé_°l (ļāëÄ-PROTAC)ŖŦšPÕßÕJéģųĶÚëÄļÎĩÄ PROTAC ŋÉÄÜĘĮÔíŧŧĐg°lÕšĩÄŌģˇŊĪōŖēÄŋĮ°÷Ŋy PROTAC (ĐĄˇÖ×ĶéÖ÷) ĐčŌĒĮ°ÆÚÅäķwĩÄЧęPĪĩŅĐžŋŌÔ´_ļ¨ģųĩAQļ¨ßBŊĶÎģÖÃŖŦĩĢÎŧĶÃĩ°°×ąžÉíĩÄļāëÄÅäķwt˛ģĐčŌĒß@ŖŦ´ķļāĩļāëÄŋÉßBŊĶĩÄÎģÖÃÔÚ C ļËģōÕß N ļËĄŖ
ŽČģŖŦČôĘĮÄî^ÔOĶ PROTAC (]ĶĐŗÉĘėÅäķwķĩĀĩÄ°Đĩ°°×)ŖŦŌĒŋŧ]ĩĞͲģHHĘĮÉĪĘöî}ÁËŖŦŊYēĪĮģĄĸÍģץĸÆäËü×ÎģücĄĸŋÉÄæš˛rŊYēĪĩȡNˇNŌōËØļŧĐčŌĒÉîČëŅĐžŋŖŦŋŊYÆđítĘĮŌģËģ¯Õnî}ĩÄķwÁŋĄŖ
Ąö E3 ßBŊĶøÅäķw
Ę×ĪČŖŦíŋ´ E3 ßBŊĶøÅäķwŖŦĻĶÚ E3 ßBŊĶøÅäķwÄŋĮ°ŗŖŌĘšĶÃĩÄßĘĮ CRBN ēÍ VHL ɡNÅäķwĄŖÄÁ÷ŗĖÉĪíÖvŖŦÔtÉĪĐčŌĒͨß^ E3 ßBŊĶøԊēĐĩÄ´_ļ¨°Đŧ°ûÖĐ E3 ßBŊĶøĩÄØSļČŖŦÄļøßxņ×îŧŅĩÄ E3 ßBŊĶøÅäķwĄŖĩĢÄŋĮ°ß@ŌģKÄŋĮ°ĩÄŅĐžŋÉæŧ°ČÔČģ˛ģĘĮēÜļāŖŦŌģ°ãļøŅÔŖŦÉĖIģ¯ĩÄÔOĶÖģĘĮßxņŊúŌ×ĩÃĩÄ CRBN ēÍ VHL ÅäķwˇÖ×ĶĄŖ

D 3. ŗŖŌĩÄ E3 ßBŊĶøŧ°ÆäÅäķw[6]
ëSÖøĻ PROTAC ŧŧĐgĩÄÉîČëŅĐžŋŖŦÄŋĮ°Ļ E3 ˇēËØßBŊĶøÅäķwĩÄēYßxŌ˛é_ĘŧßMČëŋÆWŧŌŌŌ°ĄŖÔÚ “ČũÔĒŊĩŊâÍēĪˇÖ×Ķ” ĩÄÄŖĘŊĪÂŖŦŌ˛ŗöŦFÁËąČČįģųĶÚ RNA øĩÄ RIBOTAC (ÆäšĻÄÜĘĮĸÄŋË RNA “§ĩŊ” RNaseßMĐĐŊĩŊâ)ĄĸģųĶÚČÜøķwĩÄ ATTEC (ŧČŋÉŌÔÅc×ÔĘÉß^ŗĖÖĐĩÄęPæIĩ°°× LC3 ŊYēĪŖŦŌ˛ŋÉŌÔÅc°Đĩ°°×ŊYēĪŖŦĸ°Đĩ°°×Ŋļ¨ÖÁ×ÔĘÉĐĄķwßMĐĐŊĩŊâ)ĄĸLYTACŖ¨¸ÉØØËŊįĐ—— LYTAC Åc°ĐĪōĩ°°×ŊĩŊâŧŧĐg) ēÍ×ÔĘÉŊé§ĩÄŊĩŊ⊠AUTAC ĩČŖŦËüŅaŗäÁË PROTAC ĩÄŊĩŊâˇļúŖŦĘš°ĐĪōŊĩŊâŋÉŌÔ×÷ĶÃĶÚ¸üļāˇŊÃæĄŖŊYÉĪÔOĶËŧ¡˛ģ×ŖŦÖģĘĮĻ E3 ßBŊĶøÅäķw×öĪāĒĩÄĖæQĄŖ
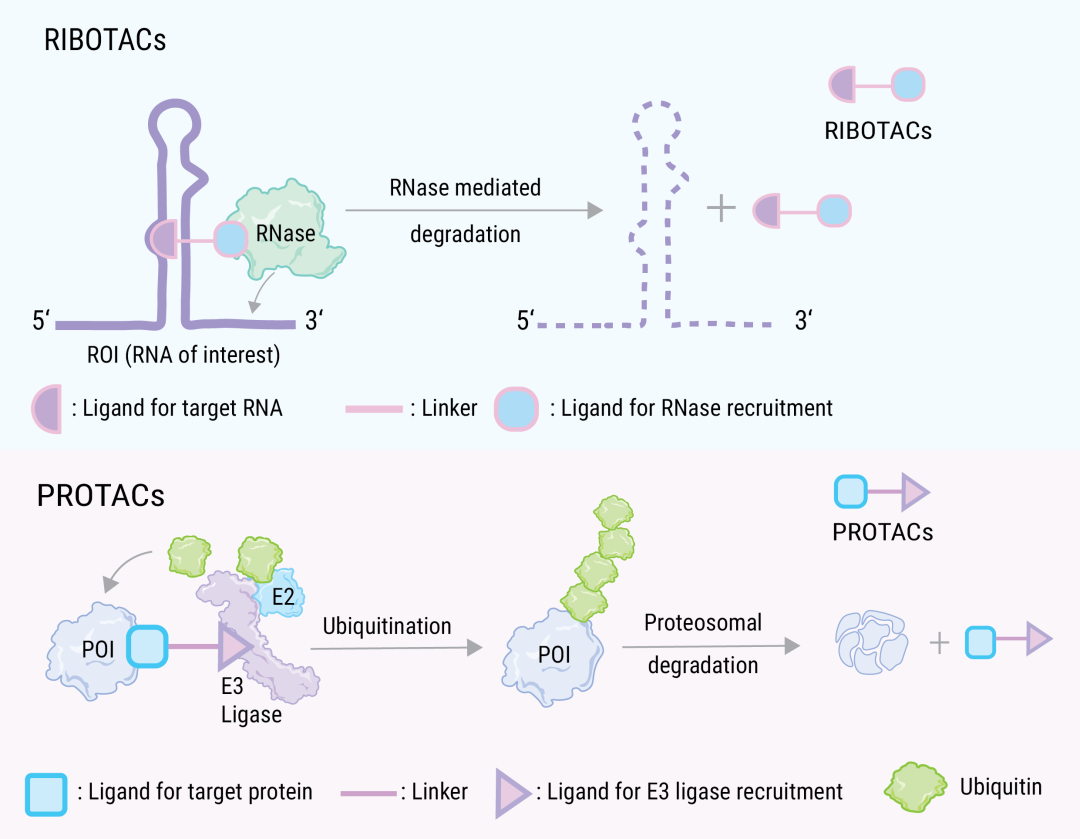
D 4. ĀûĶà RNA ø°ĐĪōŊĩŊâ ROI ĩÄ RIBOTAC[7]
a. RIBOTAC ĘĮŌģļūrˇÖ×ĶŖŦ°üēŦ RNA ŊYēĪÄŖK (×ĪÉĢ)ŖŦēËĖĮēËËáø (RNase) ÕĐÄŧÄŖK(Ë{ÉĢ)ēÍŌģßBŊĶ×Ķ (ˇÛž); RIBOTACŌģĩŠÅc°Đ RNA ŊYēĪŖŦžÍūÔÚ°Đ RNA ¸ŊŊüÕĐÄŧ RNaseŖŦÄļø´ŲßMÆäŊĩŊâĄŖb. PROTAC ŊĩŊâCÖÆĄŖ
Ąö Linker ˇÖ×Ķ
×îēķ×ÎŌŋ´ŋ´ŋ´ËÆēÎĩÄ Linker ˇÖ×ĶĄŖŗõ˛ŊĩÄ PROTAC ÔOĶŗŖŗŖūßxņ PEG æ×÷éßBŊĶˇÖ×ĶĄŖPEG æžßĶĐ×ãōĩÄČáĐÔŖŦŋÉŌÔÕ{ÕûˇÖ×ĶĩŊßmŌËĩÄŊYēĪŊĮļČŖŦĩĢŌ˛ÕũĶÉĶÚ PEG æĩÄČáĐÔŖŦסÖ×ĶĩÄŊYēĪėØß^¸ßŖŦß@ŌģücÔÚēķÆÚĩÄģ¯ÖĐĐčŌĒĸÖŽ¸ÄÉÆĄŖ
Linker ĩÄÔOĶÄŋĮ°]ĶĐŗÉĘėĩġŊ°¸ŖŦͨß^ĻÄŋĮ°ķĩĀēÍÅR´˛ŌŅÖĒĩÄ PROTAC ŊyĶŖŦ´ķ˛ŋˇÖ Linker ËųĶÃÔ×ĶĩÄæéLŗŖÔÚ 10~20 Ô×ĶéLļČĄŖÁíÍâŌ˛ĶĐŅĐžŋͨß^wŌģģ¯ˇÖÎöŖŦŋŊYÁËæéLÅcŊĩŊâģîĐÔĩÄęPĪĩŖŦÄĪÂDÖĐ´ķÖÂŋÉŌÔŋ´ŗöŖēŨ^éLĩÄæéLĶÉĶÚ¸ü¸ßĩÄėØļøĘšģîĐÔŊĩĩÍŖŦĩĢĪāŨ^ĶÚŨ^ĩÍæéL§ÖÂĩÄoˇ¨ĶĐЧĀŊüÅäķwÉļˡÖ×Ķžāëxß@ŌģŊYšûŖŦËüĩÄĶ°íŨ^ĐĄĄŖËųŌÔˇÖ×ĶÔOĶŗõÆÚŌģ°ãūßxņÉÔéLĩÄ Linker æĄŖ
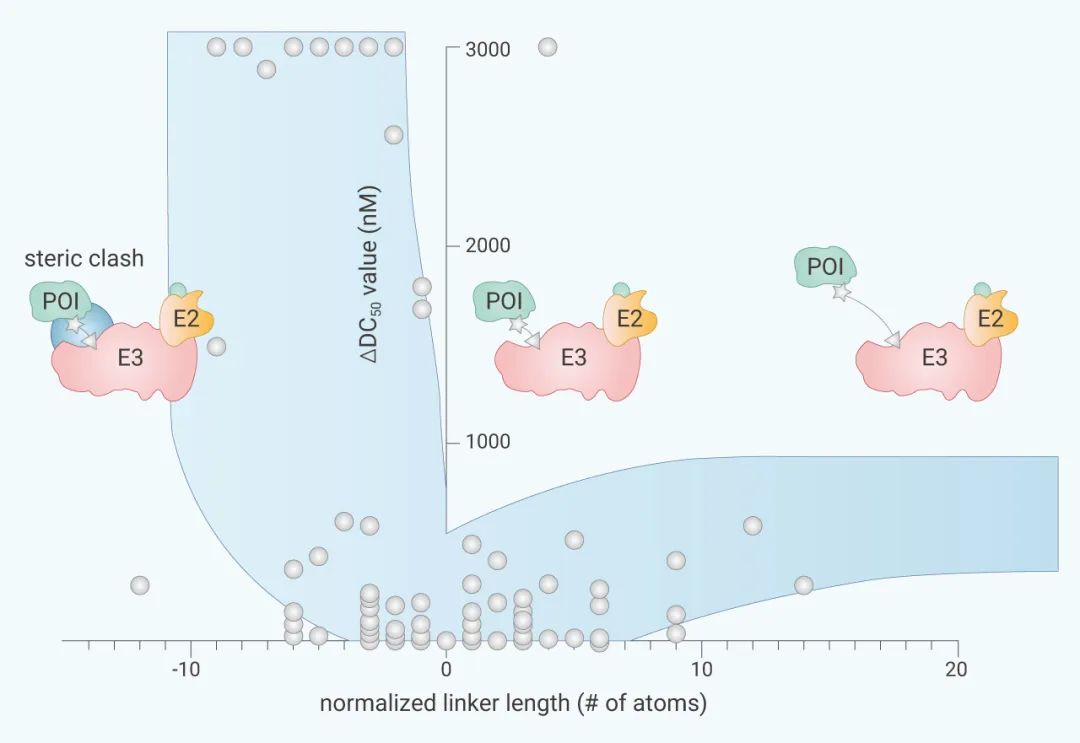
žßķwî}ĐčŌĒžßķwˇÖÎöŖŦLinker ĩÄßxņ˛ģĩĢÅcĩ°°×ŊYēĪĶōĩÄÉîļČĪāęPŖŦŌ˛ūĻÅäķwÅcĩ°°×ĩÄŊYēĪŌÔŧ°Õûķw PROTAC ĩÄÎīĀíĐÔŲ|ÔėŗÉĶ°íĄŖÔÚ PROTAC ĩÄēķÆÚģ¯ÖĐŖŦËüÍųÍųĘĮŋŧ˛ėĩÄÖØücĄŖ
Õ\ČģŖŦŊß^ŌÔÉĪײŊēĪĀíÔOĶēķëmČģŋÉŌÔŗõ˛ŊŊ¨Ōģ PROTAC ˇÖ×ĶŖŦĩĢËü˛ĸ˛ģŌģļ¨žßĶĐģîĐÔŖŦČÎēÎËÎīĩÄÔOĶļŧ˛ģĘĮŌģõíļøžÍŖŦļøĘĮōō×CÅcŋŊYÔOĶÂŨĐũÉĪÉũĩÄß^ŗĖĄŖÔÚēķÆÚĩÄŊYģ¯ˇÖÎöÔŌōrŖŦČÔČģĐčŌĒžCēĪŋŧÁŋÃŋŌģ˛ŊÔOĶÖĐßxņĩġÖ×ĶĄĸßBŊĶÎģücĩÄēĪĀíĐÔŌÔŧ°Õûķw PROTAC ĩÄ͸ĤĐÔĩČî}ĄŖ
MCE íĶĐØS¸ģĩÄ PROTAC ŗÉÆˇˇÖ×ĶŖŦÉîĩÃēŖČÍâąļāŋÆŅĐĶÃôĩÄĮā˛AĄŖ´ËÍâŖŦÎŌßĖᚊŖIĩÄ PROTAC ÔOĶ×ÉÔˇūÕŖŦÖúÁĻÄú¸ßЧēĪĀíËÎīÔOĶĄŖÎŌíĶĐŊōØS¸ģĩÄēĪŗÉŧŧĐgFę ŖŦÔÚ×îŊK PROTAC ˇÖ×ĶÉĪéÄúĖᚊŋėËŲĄĸŊúĄĸ¸ßŲ|ÁŋĩÄēĪŗɡūÕĄŖ


| PROTAC |
|
ĘĮŌģˇN¸ßЧĩÄŖŦßxņĐÔĩÄŖŦŧ°û͸ß^ĩÄģųĶÚ PROTAC ŧŧĐgĩÄ BET ŊĩŊâŠŖŦIC50 Öĩé 14 nMŖŦžßĶĐŋšÄ[ÁöģîĐÔĄŖ |
|
ĘĮŌģˇNģųĶÚ PROTAC ŧŧĐgĩÄĶĐЧĩÄ K-Ras ŊĩŊâŠŖŦÔÚ SW1573 ÖĐĻ K-Ras ŊĩŊâÂĘ ≥ 70%ĄŖ |
| SNIPER |
|
ĶÉ IAP Ū×ŋšŠ LCL-161 ŅÜÉúÎīēÍ BET ŌÖÖÆŠ (+)-JQ-1ŖŦͨß^ linker ßBŊĶŊMŗÉŖŦÕT§ BRD4 ŊĩŊâĄŖSNIPER (BRD)-1 ÍŦrŌÖÖÆ cIAP1ŖŦcIAP2 ēÍ XIAPŖŦIC50 ˇÖeé 6.8 nMŖŦ17 nM ēÍ 49 nMĄŖ |
|
ĶÉ Dasatinib (ABL ŌÖÖÆŠ) ͨß^ linker Åc Bestatin (cIAP1 Åäķw) ŊMēĪļøŗÉŖŦŋÉĶĐЧŊĩŊâ BCR-ABL ĩ°°×ĄŖ |
| Protac-linker conjugate for PAC |
|
PROTAC BRD4 Degrader-5-CO-PEG3-N3 ĘĮŌģˇNĶÃĶÚ PAC ĩÄ PROTAC-linker ÅŧÂÎīŖŦ°üēŦ BRD4 ŊĩŊ⊠GNE-987 ēÍ 3 PEG ĩÄ linkerĄŖ |
|
ĶÉ ADC linker ēÍ PROTAC ˇÖ×ĶŊMŗÉŖŦPAC ÅcŋšķwÅŧÂĄŖÅc PROTAC (˛ģÅŧ Ab) ĪāąČŖŦPAC ÅŧÂŋšķwÖŽēķ¸üŧĶī@ÖøŊĩĩÍ´Æŧ¤ËØĘÜķw-α (ERα) ËŽÆŊĄŖ |
|
PROTAC BRD4 degrader for PAC-1 ĘĮŌģˇNĶÃĶÚ PAC ĩÄ PROTAC-linker ÅŧÂÎīŖŦ°üēŦĮļēĪķw BET ŊĩŊ⊠GNE-987 ēÍēŦļūÁōģ¯ÎīĩÄ linkerĄŖ |
MCE ĩÄËųĶĐŽaÆˇHĶÃ×÷ŋÆWŅĐžŋģōË×CÉęķŖŦÎŌ˛ģéČÎēÎČËĶÃÍžĖᚊŽaÆˇē͡ūÕĄŖ
ĸŋŧÎÄĢI
1. Tao AJ, Gadbois GE, Buczynski SA, Ferguson FM. Targeted protein degradation: Emerging concepts and protein state-specific targeting principles. Curr Opin Chem Biol. 2022 Jan 15;67:102114.
2. Chen Y, Yuan X, Tang M, Shi M, Yang T, Liu K, Deng D, Chen L. Degrading FLT3-ITD protein by proteolysis targeting chimera (PROTAC). Bioorg Chem. 2022 Feb;119:105508.3. Schneider M, Radoux CJ, Hercules A, Ochoa D, Dunham I, Zalmas LP, Hessler G, Ruf S, Shanmugasundaram V, Hann MM, Thomas PJ, Queisser MA, Benowitz AB, Brown K, Leach AR. The PROTACtable genome. Nat Rev Drug Discov. 2021 Jul 20; 20:789-797.
4. Jin J, Wu Y, Chen J, Shen Y, Zhang L, Zhang H, Chen L, Yuan H, Chen H, Zhang W, Luan X. The peptide PROTAC modality: a novel strategy for targeted protein ubiquitination. Theranostics. 2020 Aug 8;10(22):10141-10153.
5. Fischer F, Alves Avelar LA, Murray L, Kurz T. Designing HDAC-PROTACs: lessons learned so far. Future Med Chem. 2022 Jan;14(3):143-166.
6. Zhong Y, Chi F, Wu H, Liu Y, Xie Z, Huang W, Shi W, Qian H. Emerging targeted protein degradation tools for innovative drug discovery: From classical PROTACs to the novel and beyond. Eur J Med Chem. 2022 Jan 20;231:114142.
7. Costales MG, Suresh B, Vishnu K, Disney MD. Targeted Degradation of a Hypoxia-Associated Non-coding RNA Enhances the Selectivity of a Small Molecule Interacting with RNA. Cell Chem Biol. 2019 Aug 15;26(8):1180-1186.e5.
8. Bemis TA, La Clair JJ, Burkart MD. Unraveling the Role of Linker Design in Proteolysis Targeting Chimeras. J Med Chem. 2021 Jun 24;64(12):8042-8052.

- 2025AACRÄęūAMÂäÄģŖŦMCEyÔŠŽaÆˇÁÁĪāÖĨŧĶ¸į
- Î÷ÃĀŊÜŗÉšĻ ĸŧĶĩÚĘŽÃÉúÎīÖÆˡļ¨ĐÔÕ¯
- Íļ¸åĶĐļY-2025 MCEÖĐøÉúÃüŋÆWŅĐžŋ´ŲßMĒÕũĘŊĸĶ
- °ŦĨÍØ&°˛ÍØËŧū×hŅûÕŖēŧ¯˛Ér´úĩÄÍësÖÆŠĐ¡ŊĪō
- é_WŧžŖŦŋÆŅиŖĀûĩŊ!ąžÉúÉúÎīĖáČĄÔŠēĐĪŪrĖØģŨ
- Î÷°˛°ŲÎÉúÎīĀŌ°ˇČžÁĪĪŪr´ŲäNŖŦŲŲIŧ´ÃâŲMËÍëĶ°Æą
- ÃĀĩÂÂÍÆŗöͨß^LC-MS/MSU´ķĐÂÉúēēY˛éĩÄzyÔŠ
- Î÷ÃĀŊÜÕũĘŊÍÆŗöUSP/EPËĘÆˇ˛ÉŲˇūÕ


