
ҶОјҡ°ыRNA/ө°°ЧҪMңyРтиb¶ЁРЎКуТИҚuјҡ°ыҒҶИәҢҰёЯЦ¬пӢКіҙъЦxЯm‘ӘРФХ{ҝШ

ЖЪҝҜЈәDiabetologiaЈЁIF=10.460Ј©
јјРg·ю„ХЈәҶОјҡ°ыRNAңyРтЎўҶОјҡ°ыө°°ЧҪMңyРтЈЁУЙЙПәЈІ®әАЙъОпМṩјјРg·ю„ХЈ©
Ң§ХZ
ТИҚuКЗТ»·N°ьә¬¶аоҗјҡ°ыөДҫ«ГЬОўРЎЖч№ЩЈ¬ЖдЦРαЎўβәНδөИғИ·ЦГЪјҡ°ы…ўЕcҷCуwСӘМЗ·Җ‘BөДХ{ҝШЎЈҙуБҝСРҫҝұнГчЈ¬·КЕЦХTҢ§өД2РНМЗДтІЎФзЖЪјҙіц¬FТИҚuЛШөЦҝ№°йлSТИҚu№ҰДЬХПөKЈ¬ТИҚuαЎўβЎўδјҡ°ыјӨЛШ·ЦГЪй_КјОЙҒyЈ¬лSЦшТИҚu№ҰДЬ“pӮыөДјУ„ЎЈ¬»јХЯІЎЗйТІЦрқuҗә»ҜЎЈҮ§эXоҗ„УОп»тИЛТИҚu¶јҙжФЪҸНлsөДҝХйgҪYҳӢЈ¬јҙК№ПаН¬оҗРНјҡ°ыйgТІҙжФЪ®җЩ|РФЈ¬¶шјІІЎ—lјюПВН¬·NоҗРНјҡ°ыҒҶИәИзәО°lЙъЧғ»ҜІўУ°н‘ТИҚu№ҰДЬөДҷCЦЖІўІ»Гчҙ_ЎЈТтҙЛЈ¬ХЖОХ2РНМЗДтІЎФзЖЪТИҚuαЎўβЎўδјҡ°ыҒҶИәөДҪMіЙЧғ»ҜЈ¬ІўМҪЛчҝЙУГУЪ…^·ЦёчҒҶИәөДұнГжҳЛЦҫОпЈ¬ҢҰ2РНМЗДтІЎөДҫ«ңКЦОҜҹәНоA·АҫЯУРК®·ЦЦШТӘөД¬FҢҚТвБxЎЈ
ҢҚтһ·Ҫ·Ё
ІЙУГҶОјҡ°ыRNAңyРт(scRNA-seq)·Ҫ·Ёиb¶ЁhfdХTҢ§өДРЎКуЖПМСМЗДНКЬІ»БјДЈРНЦРөДαЎўβәНδјҡ°ыҒҶИәҳЛУӣОпЎЈІЙУГБчКҪјҡ°ығxәНГвТЯИҫЙ«ҢҰёчҒҶИәөДұИАэЯMРР·ЦоҗәНФu№АЎЈҢҰ·ЦЯxјҡ°ыЯMРРҶОјҡ°ыө°°ЧЩ|ҪMҢW·ЦОцЈ¬НЁЯ^ө°°ЧұнЯ_ЙоИлкUГчёчαЎўβЎўδјҡ°ыҒҶИәФЪЖПМСМЗДНКЬІ»БјЦРөД№ҰДЬ о‘BЎЈ
СРҫҝҪY№ы
1. РЎКуТИҚuҶОјҡ°ыRNAңyРт
ұҫСРҫҝҢҰ5Ц»ЖХНЁпӢКіЈЁcdЈ©О№рBөДРЎКуәН5Ц»ёЯЦ¬пӢКіЈЁhfdЈ©О№рBөДРЎКуЯMРРБЛёЯНЁБҝscRNA-seq·ЦОцЎЈСРҫҝ·ЦОцБЛ15093ӮҖҒнЧФcdО№рBРЎКуТИҚuөДјҡ°ыәН18906ӮҖҒнЧФhfdО№рBРЎКуТИҚuөДјҡ°ыЈ¬°l¬FЖдұ»ҫЫіЙЛД·NЦчТӘғИ·ЦГЪјҡ°ыЈ¬»щУЪЦчТӘјӨЛШ»щТтұнЯ_ЯMРРБЛЧўбҢ:4069ӮҖα (Gcg)Ј¬ 14108ӮҖβ (Ins1)Ј¬ 1698ӮҖδ (Sst)әН666ӮҖPP (Ppy)јҡ°ыЎЈҙЛНвЈ¬ёщ“юПИЗ°ҲуөАөДҳЛУӣ»щТтЈ¬°l¬FБЛЙЩБҝғИЖӨјҡ°ыЎўГвТЯјҡ°ыЎўйgідЩ|јҡ°ыәНҢ§№Ьјҡ°ыЈЁҲD1-dЈ©ЎЈҪУПВҒнҢҰαЈ¬βЈ¬δЯ@ҺЧ·Nјҡ°ыоҗРНЯMРРІо®җ»щТтәН№ҰДЬё»јҜ·ЦОцЈ¬°l¬FғИЩ|ҫWөДө°°ЧЩ|јУ№ӨФЪαЎўβәНδјҡ°ыЦРп@Цшё»јҜЈЁҲD1e-gЈ©ЎЈ
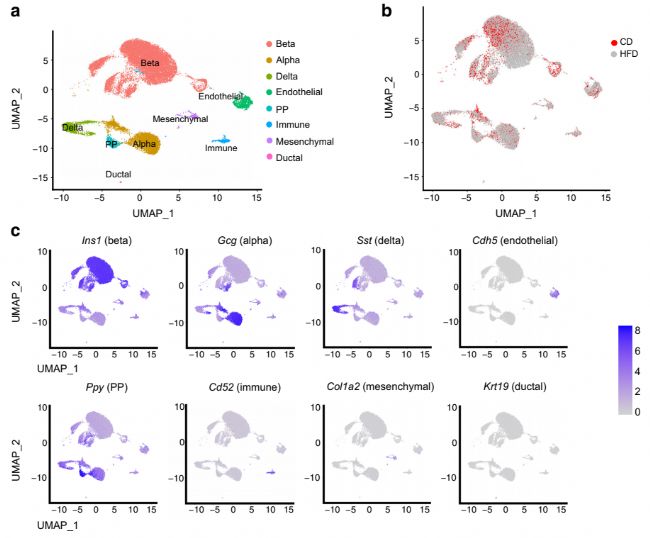
ҲD1
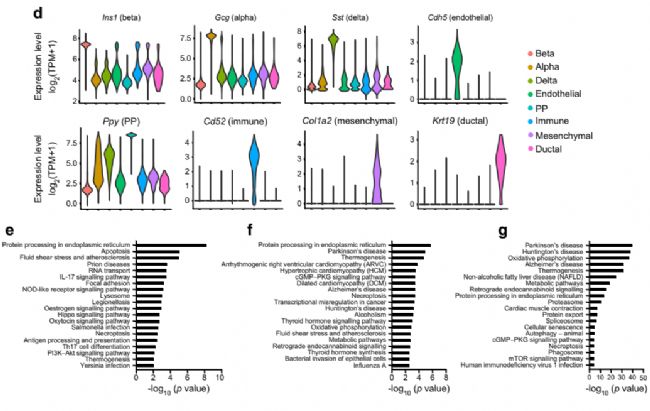
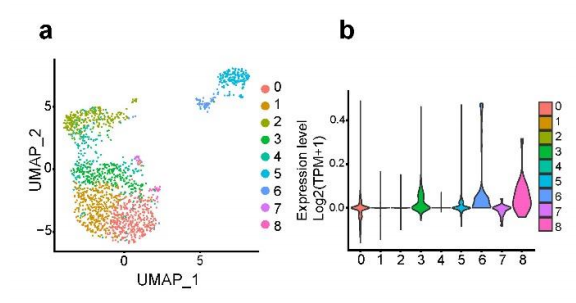
2. Ace2ФЪαјҡ°ыЦРөД®җЩ|ұнЯ_
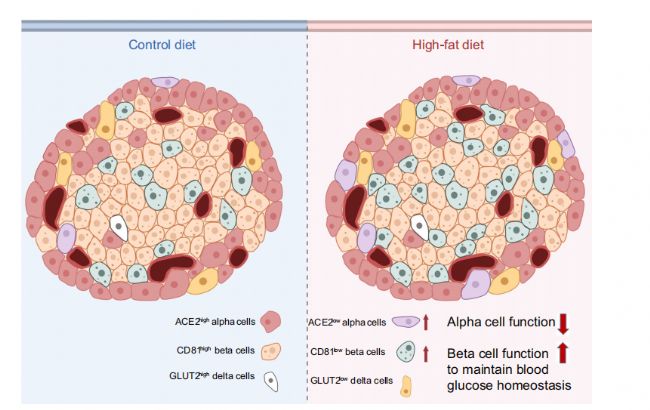
һйБЛ·ЦОцαјҡ°ыөД®җЩ|РФЈ¬К№УГscRNA-seq”ө“юҢўαјҡ°ыҫЫіЙ8ӮҖҙШЈЁҲD2a-bЈ©Ј¬Іў·ЦОцБЛ8ӮҖҫЫоҗЦРөДmarker»щТтұнЯ_ЈЁҲD2cЈ©ЎЈұҫСРҫҝ°l¬FЕcҮАЦШјұРФәфОьҫCәПХч№Ъ оІЎ¶ҫ2 (SARS-CoV-2)ёРИҫПакPөДкPжI»щТтAce2ФЪαјҡ°ыИәЦРп@ЦшЧғ»ҜЎЈёщ“юAce2өДұнЯ_Л®ЖҪЈ¬Ңў8ӮҖαјҡ°ыҙШ·ЦһйAce2low(ҙШ3әН5)әНAce2high(ҙШ0Ўў1Ўў2Ўў4Ўў6әН7)ҒҶИәЎЈScRNA-seqәНГвТЯҹЙ№вЧCҢҚБЛФЪcdәНhfdРЎКуТИҚuЦРαјҡ°ыҒҶИәөДҙжФЪЈЁҲDf,g,jЈ©ЎЈһйБЛМҪЛчace2 αјҡ°ы®җЩ|РФөД·ЦЧУМШХчЈ¬ОТӮғФЪAce2lowәНAce2highҒҶИәЦ®йgиb¶Ёіц379ӮҖІо®җ»щТтЈЁҲD2hЈ©Ј¬ІўҢҰІо®җ»щТтЯMРРБЛKEGGё»јҜ·ЦОцЈЁҲD2iЈ©Ј¬ұнГчAce2өДұнЯ_ҝЙТФ…^·ЦҫЯУРІ»Н¬№ҰДЬәНЙъҙж о‘BөДαјҡ°ыЎЈОТӮғЯҖ°l¬FЈ¬ФЪAce2lowҪMЦРЈ¬αјҡ°ыМШ®җРФЮDдӣТтЧУЈЁMafbәНIrx1Ј©әНαјҡ°ыПакP»щТтЈЁTtrЎўChgbЎўFevәНPyyЈ©п@ЦшПВХ{ЎЈПа·ҙЈ¬Ace2low αјҡ°ый_КјұнЯ_ҫЯУРβјҡ°ыМШХчөД»щТтЈЁИзUcn3ЎўEro1lbЎўәНSlc2a2Ј©ЎЈЖдЦРТ»Р©ҪY№ыНЁЯ^qPCRЈЁҲD2m-qЈ©әНwesternУЎЫEЈЁҲD2kЈ©ҢҰ·ЦЯxөДαјҡ°ыИәЯMРРБЛЧCҢҚЎЈЯ@ұнГчЈ¬Ace2lowҒҶИәҝЙДЬҙъұнБЛОҙіЙКмөДαјҡ°ыЈ¬ЛЖәхҫЯУРβјҡ°ыҳУМШХчЎЈ


ҲD2
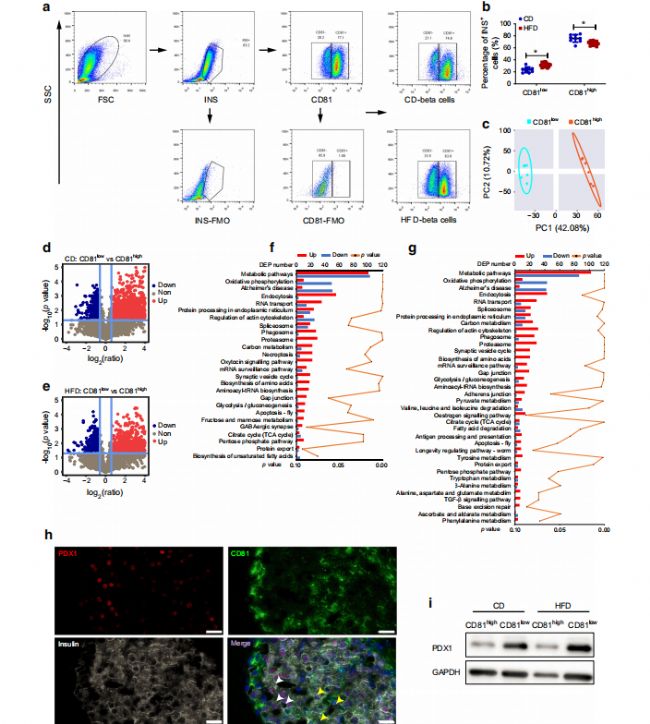
3. ұнГчαјҡ°ыҒҶИә№ҰДЬөДҶОјҡ°ыө°°ЧЩ|ҪMҢW
УГТИҚuҶОјҡ°ы‘ТТәөД FACSҢўαјҡ°ыЈЁТИёЯСӘМЗЛШ[GCG]+Ј©јҡ°ы·ЦһйACE2highҪMәНACE2lowҪMЎЈЦөөГЧўТвөДКЗЈ¬HFDРЎКуЦРACE2high αјҡ°ыөДұИАэЈЁ65.3%Ј©Гчп@өНУЪCDРЎКуЈЁ80.0%Ј©,Я@ұнГч»щУЪace2өДαјҡ°ы®җЩ|РФҝЙДЬ…ўЕcБЛМЗДтІЎөДІЎАнЙъАнЯ^іМЈЁҲD3a,bЈ©ЎЈҸДFACS·ЦЯxөДөДCDәНHFDРЎКуЈЁГҝӮҖҳУұҫ2000ӮҖјҡ°ыЈ©ЦРКХјҜACE2lowәНACE2high αјҡ°ыҒҶИәЯMРРҶОјҡ°ыө°°ЧЩ|ҪMҢW·ЦОцЎЈҷzңyөҪөДө°°Ч”өБҝФЪ3042~4021Ц®йgЈ¬ЦШҸНРФБјәГЎЈPCAТІДЬГчп@…^·ЦACE2lowјҡ°ыәНACE2highјҡ°ыЈЁҲD3cЈ©ЎЈФЪCDРЎКуЦРЈ¬ЕcACE2high αјҡ°ыПаұИЈ¬ACE2low αјҡ°ыЦРУР29ӮҖЙПХ{Ј¬80ӮҖDEPsПВХ{(ҲD3d)Ј¬КЬУ°н‘өДKEGGНЁВ·°ьАЁRasРЕМ–НЁВ·әНУРҪz·ЦБС»о»Ҝө°°ЧјӨГёЈЁMAPKЈ©РЕМ–НЁВ·ЈЁҲD3 eЈ©ЎЈФЪHFDРЎКуЦРЈ¬ЕcACE2high αјҡ°ыПаұИЈ¬ACE2low αјҡ°ыЦРөДDEPsГчп@ФцјУЈ¬ЖдЦР17ӮҖЙПХ{Ј¬301ӮҖDEPsПВХ{ЈЁҲD3 fЈ©ЎЈKEGG·ЦОц°l¬FЕc IрBәНДЬБҝҙъЦxПакPөДНҫҸҪФЪACE2өНҪMҫщп@ЦшПВХ{ЈЁҲD3gЈ©ЎЈЯ@Р©°l¬FҸҠБТ°өКҫБЛACE2low αјҡ°ыөДҙъЦxәНЖдЛыЙъОп»оРФңpИхЈ¬МШ„eКЗФЪHFDРЎКуЦРЎЈ
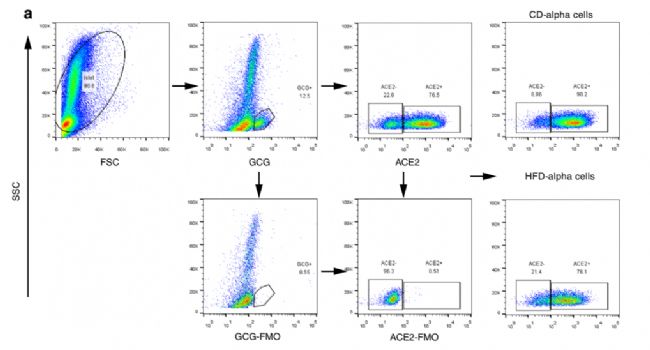

ҲD3
4. CD81ФЪβјҡ°ыЦРөД®җЩ|ұнЯ_
һйБЛ·ЦОцβјҡ°ыөД®җЩ|РФЈ¬ОТӮғҢўЯ@Р©јҡ°ыҫЫһй9ӮҖҙШЈ¬Іўҙ_¶ЁБЛЗ°10ӮҖҳЛУӣ»щТтЈЁҲD4a-cЈ©ЎЈёщ“юЧоҪьөДТ»н—СРҫҝЈ¬°l¬FCd81ФЪјҡ°ыҙШЦРУРІо®җұнЯ_ЎЈСРҫҝҢў9ӮҖβјҡ°ыҙШ·ЦһйCd81lowЈЁҙШ0-4әН7Ј©әНCd81highЈЁҙШ5Ўў6әН8Ј©ҒҶИәЈЁ4d-gЈ©Ј¬°l¬FCD81ФЪCDәНHFDРЎКуөДβјҡ°ыЦР·ЦІјөДГчп@®җЩ|РФЈ¬Н¬•rТІҪӣЯ^БЛГвТЯҪM»ҜөДЧCҢҚЈЁҲD4jЈ©ЎЈҪУПВҒн·ЦОцБЛCd81lowПаҢҰУЪCd81 highβјҡ°ыөДІо®җ»щТтәНKEGGНЁВ··ЦОцЈ¬јҡ°ыҙъЦxј°ЖдПакP№ҰДЬНЁВ·ТФј°Т»ПөБР‘ӘјӨПакPәНСЧ°YНЁВ·ұ»п@Цшё»јҜЎЈ¶шОҙіЙКмөДјҡ°ыҳЛУӣОпЈЁChgaЎўChgbЎўMafbәНRbp4Ј©ФЪCd81highҒҶИәЦРёЯұнЯ_ЈЁҲD4lЈ©ЎЈЯ@ұнГчCd81highҒҶИәҝЙДЬҙъұнБЛТ»ҪM№ҰДЬЭ^ИхәНОҙіЙКмөДјҡ°ыЎЈЖдЦРТ»Р©ҪY№ыНЁЯ^qPCRЈЁҲD4m-pЈ©әНwesternblotЈЁҲD4kЈ©ҢҰ·ЦЯxөДјҡ°ыҒҶИәЯMРРЧCҢҚЎЈ
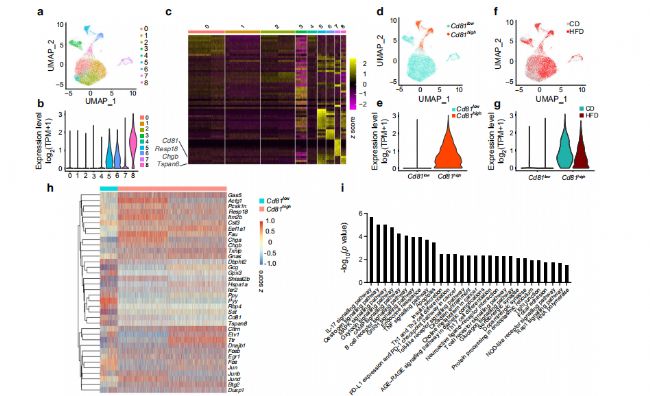

ҲD4
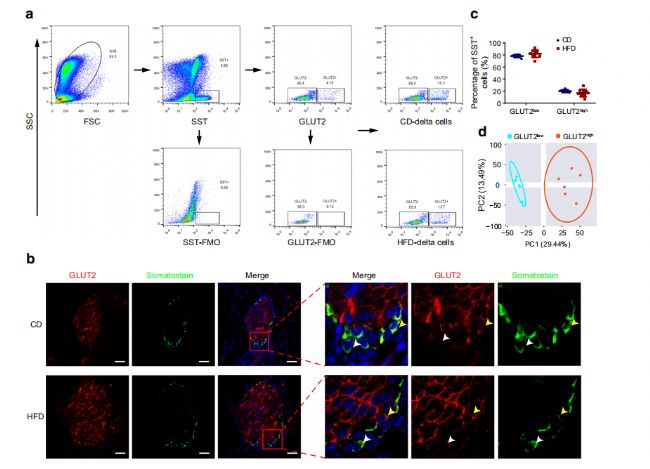
5. βјҡ°ыҒҶИә№ҰДЬөДҶОјҡ°ыө°°ЧЩ|ҪMҢWиb¶Ё
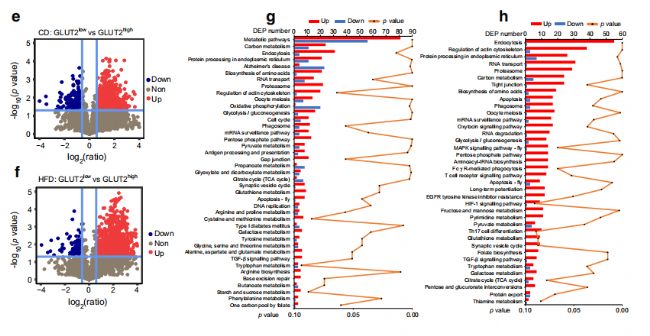
һйБЛЯMТ»ІҪМҪЛчHFDҢҰβјҡ°ыҒҶИәөДУ°н‘Ј¬FACSп@КҫЕcCDРЎКуПаұИЈ¬HFDРЎКуөДCD81highјҡ°ыұИАэҪөөНЎЈИ»әуЈ¬К№УГҶОјҡ°ыө°°ЧЩ|ҪMҢWЈЁГҝӮҖҳУұҫ10000ӮҖјҡ°ыЈ©·ЦОцFACS·ЦЯxөДCD81lowәНCD81highβјҡ°ыЈЁҲD5a,bЈ©ЎЈPCAп@КҫCD81lowәНCD81highβјҡ°ыұ»ҫЫјҜіЙғЙӮҖӘҡБўөДҪMЈЁҲD5cЈ©ЎЈұИЭ^CD81lowәНCD81highβјҡ°ыЈ¬ФЪCDРЎКуЦРҷzңyөҪ869ӮҖЙПХ{әН373ӮҖПВХ{өДDEPsЈ¬ФЪHFDРЎКуЦРҷzңyөҪ707ӮҖЙПХ{әН312ӮҖПВХ{өДDEPsЈЁҲD5d,eЈ©ЎЈФЪCDәНHFDРЎКуЦРЈ¬ЕcДЬБҝҙъЦxПакPөДНҫҸҪЈЁ°ьАЁСх»ҜБЧЛб»ҜәН°ў –ҙДәЈД¬ІЎЈ©ФЪCD81өНҒҶИәЦРұ»ПВХ{ЈЁҲD5f,gЈ©Ј¬ІўНЁЯ^ГвТЯҪM»ҜәНwestern blotЯMРРБЛтһЧCЎЈ
ҲD5
6. HFDФцҸҠБЛіЙКмөДSlc2a2 low δјҡ°ыөД№ҰДЬ
ұҫСРҫҝҢўδјҡ°ы·Цһй Slc2a2low and Slc2a2high ҒҶИәЈ¬НЁЯ^Slc2a2low ПаұИУЪ Slc2a2highІо®җ»щТтәНKEGG№ҰДЬё»јҜ·ЦОцп@КҫЈ¬ЙПХ{өДНЁВ·Еcв}лxЧУәНјӨЛШ·ЦГЪУРкPЈ¬МбКҫSlc2a2lowҒҶҪMҝЙДЬ№ҰДЬёьіЙКмЈЁS6a,bЈ©ЎЈИ»¶шЈ¬8ЦЬөДHFDІўӣ]УРёДЧғғЙӮҖdeltaјҡ°ыҒҶИәөДұИАэЈЁҲD6a,cЈ©ЎЈГвТЯҹЙ№вЧCҢҚБЛФЪРЎКуәНИЛоҗТИҚuЦРҫщҙжФЪδјҡ°ыҒҶИәЈЁҲD6bЈ©ЎЈҪУПВҒнІЙУГFASCҢҰCDәНHFDРЎКуөДSlc2a2low and Slc2a2high δјҡ°ыҒҶИәЈЁГҝӮҖҳУұҫ3000ӮҖјҡ°ыЈ©ЯMРРҶОјҡ°ыө°°ЧЩ|ҪMҢW·ЦОцЎЈPCA”ө“юп@КҫЈ¬Slc2a2low әНSlc2a2high δјҡ°ыҫЫјҜіЙғЙӮҖІ»Н¬өДИәуwЈЁҲD6dЈ©ЎЈSlc2a2low әН Slc2a2high δјҡ°ыҒҶИәЦ®йgУР548ӮҖЙПХ{әН171ӮҖПВХ{өДDEPsЈ»Н¬•rЈ¬HFDРЎКуЦРDEPsЙПХ{Э^¶аЈЁ832Ј©Ј¬ПВХ{Э^ЙЩЈЁ102Ј©ЈЁҲD6e,f)ЎЈKEGG·ЦОцп@КҫЈ¬Еcβјҡ°ыПаЛЖЈ¬CDРЎКуөДSlc2a2low ЈЁПаҢҰУЪSlc2a2high δјҡ°ыЈ©ЦРөДСх»ҜБЧЛб»ҜәН°ў –ҙДәЈД¬ІЎНЁВ·п@ЦшПВХ{(ҲD6g)ЎЈФЪCDәНHFDРЎКуөДSlc2a2lowјҡ°ыЦРЈ¬¶а·N IрBҙъЦxНҫҸҪТІп@ЦшЙПХ{ЎЈҫCЙПЛщКцЈ¬Slc2a2low δјҡ°ыҝЙДЬұИSlc2a2high δёьҫЯУР№ҰДЬәНҙъЦx»оРФЈ¬¶шHFDЯMТ»ІҪҙМјӨSlc2a2low δјҡ°ыөД№ҰДЬәНҙъЦxЎЈ
ҲD6
ҝӮЦ®Ј¬ОТӮғ°l¬FОҙіЙКмөДТИҚuјҡ°ыІҝ·ЦК§ИҘБЛЛьӮғөДЙн·ЭҫSіЦ»щТтЈ¬Іўй_Кјұн¬FіцЖдЛығИ·ЦГЪјҡ°ыөДМШХчЎЈФЪHFDХTҢ§өДЖПМСМЗДНКЬІ»БјЦРЈ¬өНИЭБҝәНОҙіЙКмөДACE2lowјҡ°ыөДұИАэлSЦш№ҰДЬ»оРФәНіЙКмөДCD81lowјҡ°ыұИАэөДФцјУ¶шФцјУЎЈіЙКмөДSlc2a2low δјҡ°ы№ҰДЬФцҸҠЈ¬ө«ұИАэІ»ЧғЎЈЯ@Р©„У‘BЧғ»ҜҪТКҫБЛТИҚuҢҰHFD‘ӘјӨ·ҙ‘ӘөДҙъЦxҝЙЛЬРФЎЈЯ@·NҝЙЛЬРФЯm‘ӘҝЙТФНЁЯ^ТЦЦЖёЯСӘМЗҒнөЦПыҙъЦx‘ӘјӨЈ¬»ЦҸНЖПМСМЗ·Җ‘BЈ¬ФцҸҠјҡ°ыөДөНСӘМЗЧчУГЈ¬ІўФцјУδҢҰαәНβјҡ°ыөДХ{№қЎЈ
ҲD7
…ўҝјОД«IЈә
Fu Q, Jiang H, Qian Y, et al. Single-cell RNA sequencing combined with single-cell proteomics identifies the metabolic adaptation of islet cell subpopulations to high-fat diet in mice[J]. Diabetologia, 2022: 1-17.

- Target-BSјјРgҪТКҫМЗДтІЎТэ°lІӘЖр№ҰДЬХПөKөДDNAјЧ»щ»ҜХ{ҝШҷCЦЖ
- әПіЙЙъОпҢWөЧұPјҡ°ыЦ®®…іаҪНДёіЈУГұнЯ_Эdуwј°»щТтёДФмјјРg
- Йо¶ИҪвОцCRISPRОДҺмәYЯxБчіМј°‘ӘУГ°ёАэ
- ҝЙңШәН—lјюПВёЯ·ЦұжВКҷzңym6AРЮп—өДРВm6AңyРт·Ҫ·Ёй_°lСРҫҝ
- AIИЛ№ӨЦЗДЬФЪө°°ЧЩ|ҪYҳӢоAңyЎў№ҰДЬоAңyј°ФOУӢЦРөДЧчУГ
- m5C MeRIP-seqөИҪТКҫm5CРЮп—ФЪ°©°YДНЛҺЦРөДкPжIХ{ҝШҷCЦЖЦРөД‘ӘУГ
- ChIPјјРgҪТКҫNURR1ФЪЗ°БРПЩ°©ҸД»щТтЮDдӣөҪД[БцЯMХ№ЦРөДХ{ҝШҷCЦЖ
- ғЙ·NЮDдӣТтЧУФЪҪйҢ§·¬ЗС№ыҢҚіЙКмөДұнУ^ЯzӮчХ{ҝШЦР°l“]өДкPжIЧчУГ
- 2025І®әАЙъОпҙәјҫИ«ҮшСІЦvй_ҶўЈ¬ФзшBҲуГы“ҢХјПИҷC
- ИAҙуЦЗФмёұҝӮІГЦРҮш…^ҝӮҪӣАнЕнҡgҡgТ»РРөҪФLІ®әАЙъОп
- І®әА¶ӯКВйLіцПҜҢЈҫ«МШРВЦРРЎЖуҳI°lХ№үСҙуХ“үҜІў°lСФ
- І®әАЙъОплp11ҝсҡgМШ»Э»о„УҒнТuЈ¬ПЮ•rПЮБҝ“Ң
- вщГАНЁөВНЖіціЙұҫғrуwтһҶОјҡ°ыңyРтЈЁFFPE»тИ«СӘЈ©
- І®әАЙъОп2024ПөБРЕаУ–°аЈЁкғОчХҫЈ©ҲуГый_Ҷў
- І®әАҶОјҡ°ыFlexДкЦРҫЮ»ЭЈ¬1.1W/ҳУұҫ»о„УғrјҙҢўҪШЦ№
- Л{ҫ°ҝЖРЕDAP-seqјјРgПакPОДХВ6ЯB°lЈ¬ҝӮIF 95.2


