
神經干細胞分類及神經干細胞移植在治療神經損傷中的應用
神經退行性疾病是影響全球數百萬人的高度致殘且最終致命的疾病,主要包括肌萎縮側索硬化癥 (Amyotrophic lateral sclerosis, ALS)、帕金森病 (Parkinson's disease, PD)、阿爾茨海默癥 (Alzheimer's disease, AD) 和亨廷頓病 (Huntington's disease, HD)。在神經退行性疾病中,特定的神經元亞群,例如多巴胺能和膽堿能神經元或運動神經元會逐漸退化,導致特定模式的神經系統功能障礙。傳統藥物治療用于延緩疾病進展,不能使功能修復或組織再生[1]。
神經干細胞 (Neural stem cells, NSCs) 在神經系統的可塑性、衰老、疾病和再生方面具有重要作用。基于體內移植的 NSCs 具有自我更新、增殖、多向分化、低免疫原性和遷移的特點,NSCs 移植的成為了極具前景的一種神經系統疾病療法,為腦損傷修復以及神經性疾病的治療帶來了希望。
神經干細胞 (NSCs) 被喻為中樞神經系統 (CNS) 的 “種子” 細胞,不同神經細胞系的產生起源于成體神經干細胞。神經干細胞命運的調控更是一個精細的過程,依賴于從表觀遺傳延伸到翻譯水平的復雜調控網絡,且涉及細胞外基質成分[2]。
成體 NSCs 可以自我更新或響應特定刺激分化為神經元、星形膠質細胞或少突膠質細胞 (圖 1),不同譜系的神經細胞功能各異。
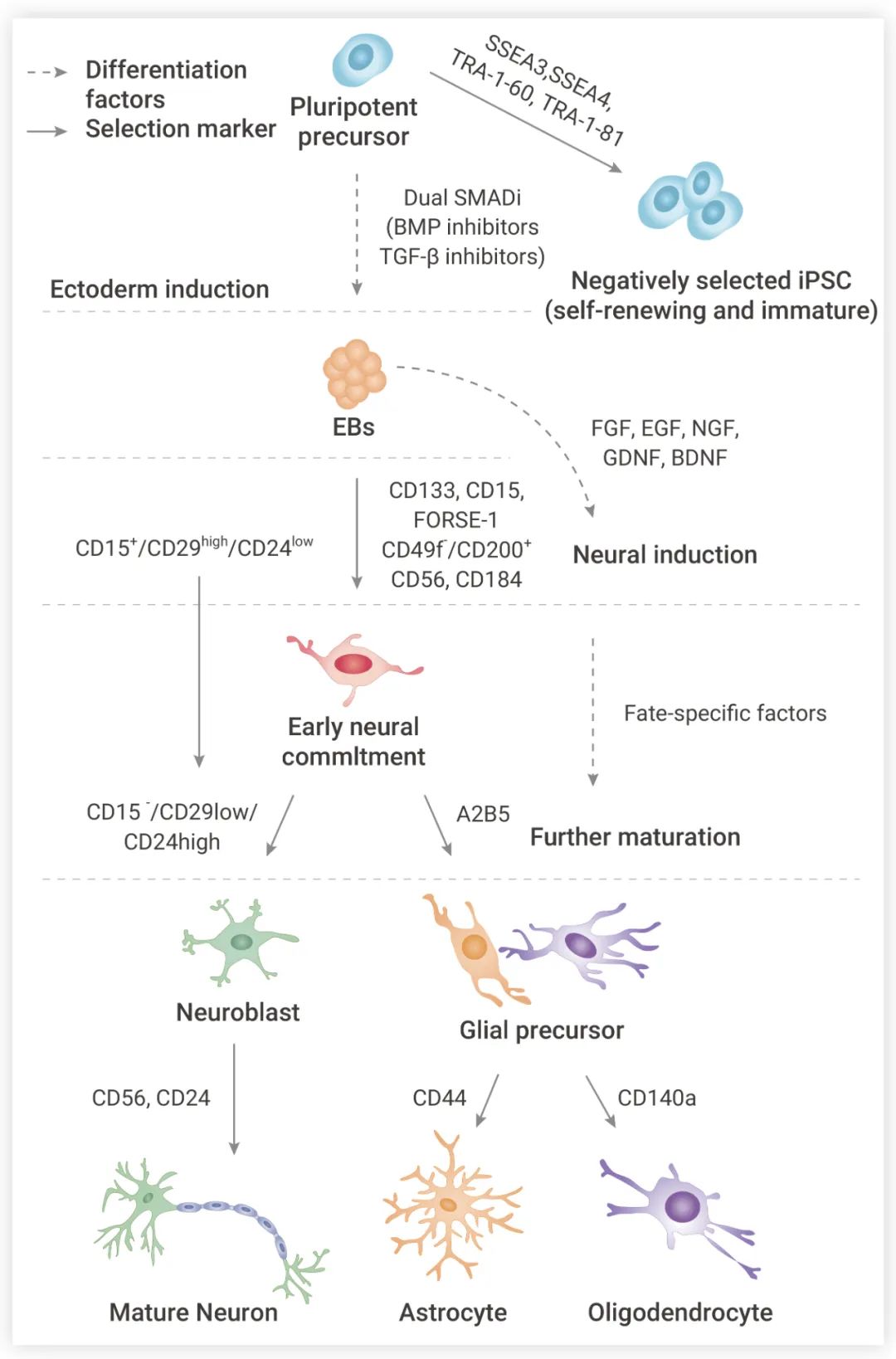
神經元 (Neurons) 是神經系統結構和功能的基本單位,通過軸突和樹突傳遞信號。典型的神經元由樹突、細胞體、軸突 (包括軸突丘) 和突觸前末端組成。神經元自身不能再生。移植外源性 NSCs 或動員內源性 NSCs,使其分化為神經元,重建損傷的神經回路,是一種被廣泛探索的治療神經退行性疾病的方法。
星形膠質細胞 (Astrocytes) 是中樞神經系統 (CNS) 中最豐富的細胞類型,它們與神經元、少突膠質細胞和內皮細胞交流,并在結構上支持它們生長,整合為三聯突觸和神經血管單位,履行其功能。除了參與細胞通訊外,星形膠質細胞還能通過釋放神經營養因子,如膠質細胞系衍生的神經營養因子 (GDNF),以及降低神經元的興奮中毒,發揮神經保護作用。
少突膠質細胞 (Oligodendrocytes, OLGs) 是有髓鞘的膠質細胞,對神經元電絕緣、促進跳躍信號傳導非常重要。少突膠質細胞還通過髓磷脂膜為神經元軸突提供代謝和營養支持,包括乳酸、丙酮酸和腦源性神經營養因子 (BDNF) 等神經營養因子。
神經干細胞,該如何培養?
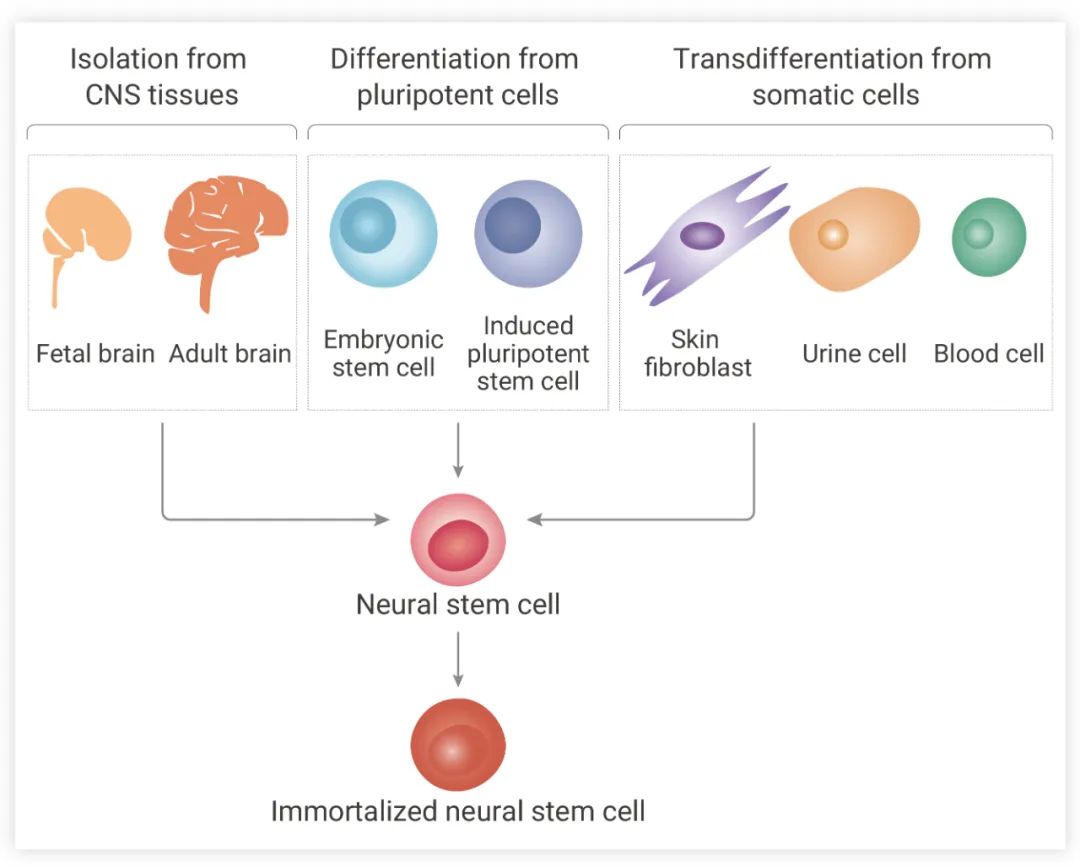
可以通過三種不同的方法獲得 NSCs (圖 2):(1) 從原發性 CNS 組織直接提取,包括腦和脊髓組織;(2) 分化多能干細胞,例如胚胎干細胞和誘導性多能干細胞;(3) 轉分化體細胞,如皮膚成纖維細胞和血細胞。從上述來源產生的 NSCs 可以通過基因改造,進一步永生化[1]。
如何有效地將 NSCs 誘導成特定的細胞類型? 神經營養因子、生長因子參與了 NSCs 增殖和分化的內源性調控,已被證明能誘導 NSCs 分化。目前已經開發了不同的 NSCs 分化方案。大多數都是基于含有生長因子的培養基,NSCs 在其中粘附和增殖。這些生長因子包括成纖維細胞生長因子 (FGF) 和/或表皮生長因子 (EGF)、神經生長因子 (NGF)、腦源性神經營養因子 (BDNF) 和膠質源性神經營養因子 (GDNF) 等。
如在下文中,作者團隊發表了一種快速、高效的分化方法,通過使用含有必要生長因子的選擇性培養基從骨髓來源的細胞中生成小鼠 OLGs。通過該方案,可以在 7-8 周內獲得成熟的 OLGs (與以前的工作相比,獲得 OLGs 的時間更短)。
作者團隊的方法主要包括三個步驟: 骨髓來源的 NSCs 生成、NSCs-OPCs 分化和 OLGs 成熟[4]。
骨髓來源的 NSCs 生成:收集骨髓組織切小塊,消化,將細胞懸浮在無血清 DMEM/F12 中,添加 2% B27 補充劑, 20 ng/mL EGF 和 10 ng/mL bFGF ,同時添加 100 IU/mL 青霉素和 100 μg/mL 鏈霉素。隨后將細胞懸浮在 NSC-PM (神經干細胞增殖培養基)中,并包裹于含聚 D-賴氨酸和層粘連蛋白的 24 孔板。10-14 天后,形成類似神經球的細胞團簇。
骨髓來源的 NSCs-OPCs 的分化和增殖:形成的神經球被消化到單細胞中,在 NSC-PM 中培養。每個孔中的神經球被消化成單細胞,在特定的 NSCs 分化培養基中培養。
星形膠質細胞分化時,DMEM 中添加 1% N2、2 mM GlutaMax-I 和 1% FBS。
OLGs 分化培養基添加 2% B27、2 mM GlutaMax-I 和 20 ng/mL T3 的 neuro 培養基。2 周后,分化培養基中的 NSCs 形態發生變化,并且能檢測到神經元、星形膠質細胞和 OLGs 的標記物。
OLGs 成熟:在 OLGs 成熟過程中,用 DPBS 緩沖液洗去生長因子 (PDGF-AA 和 bFGF),加入新鮮 EOLG-DM (早期 OLGs 分化培養基) 誘導 OLGs 分化,然后將在EOLG-DM 培養基中生長 4 天的 OPCs 轉移到 LOLG-DM (晚期 OLGs 分化培養基) 中再生長 10 天,并通過添加 cAMP 加速 OLGs 分化的分支生長。
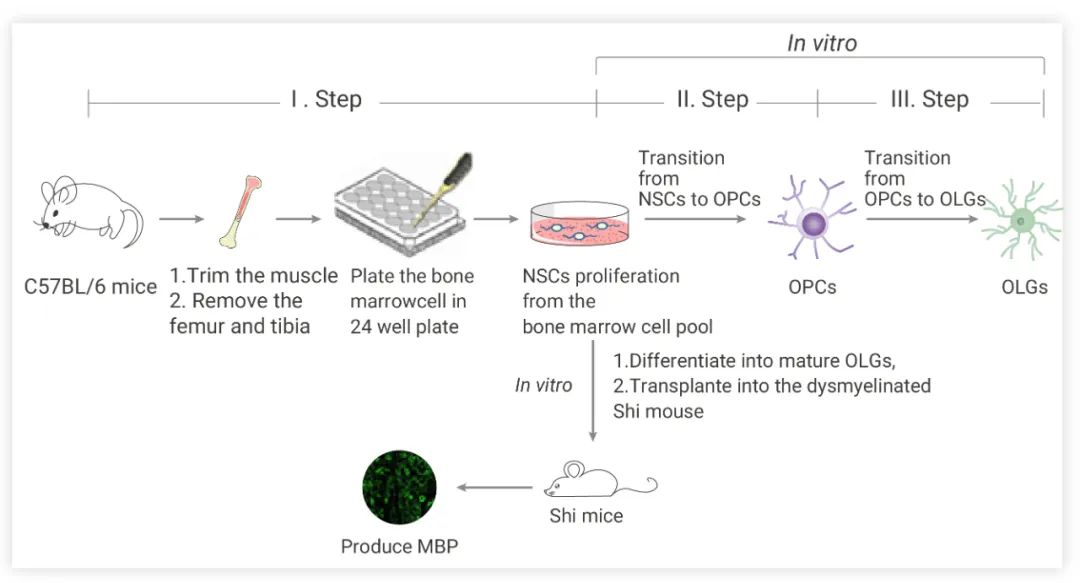
圖 3. 從小鼠自體骨髓來源的 NSCs 中生成少突細胞祖細胞 (OPCs) 和少突膠質細胞[4]
此外,越來越多的科研人員關注多種神經營養因子/細胞因子聯合使用。Kyung-Chul Choi 教授團隊證明了生長因子的組合使用 (bFGF+IGF-I、bFGF+NGF、bFGF+BDNF、BDNF+IGF-I、bFGF+NGF) 對 NSCs 分化為神經元的效果比單個生長因子 (bFGF、IGF-I、BDNF 和 NGF) 處理結果更顯著[5]。
阿爾茨海默癥 (Alzheimer's disease, AD) 是一種神經退行性疾病,由皮質和皮質下區域的神經元數量顯著減少引起,其發展主要與淀粉樣蛋白 (Aβ) 蛋白的積累和 Tau 蛋白的過度纏結相關。AD 的主要癥狀包括記憶力減退和認知障礙,大部分患者為老年人。AD 不僅降低老年人的生活質量,也加劇老齡化相關的經濟負擔,因此,AD 的治療是眾多研究者重點關注的課題。
目前治療 AD 的常用藥物有多奈哌齊、卡巴拉汀、加蘭他敏、他克林4 種乙酰膽堿酯酶抑制劑,以及 1 種 NMDA 受體拮抗劑美金剛。但藥物無法再生已損傷或丟失的神經元,對于已有明顯癥狀患者治療效果并不明顯。
NSCs 能夠自我更新并分化各種神經細胞,NSCs 移植療法已顯示出治療 AD 的巨大潛力。移植后,NSCs 分化為神經元和/或神經膠質細胞并釋放營養因子,從而修復 AD 引起的認知障礙 (圖 4)。然而,盡管 NSCs 移植能再生神經元,但對已產生的 Aβ 卻無法有效清除,這對移植后 NSCs 的存活也有不利的影響。對于 NSCs 的改造也是一個新興的研究課題。
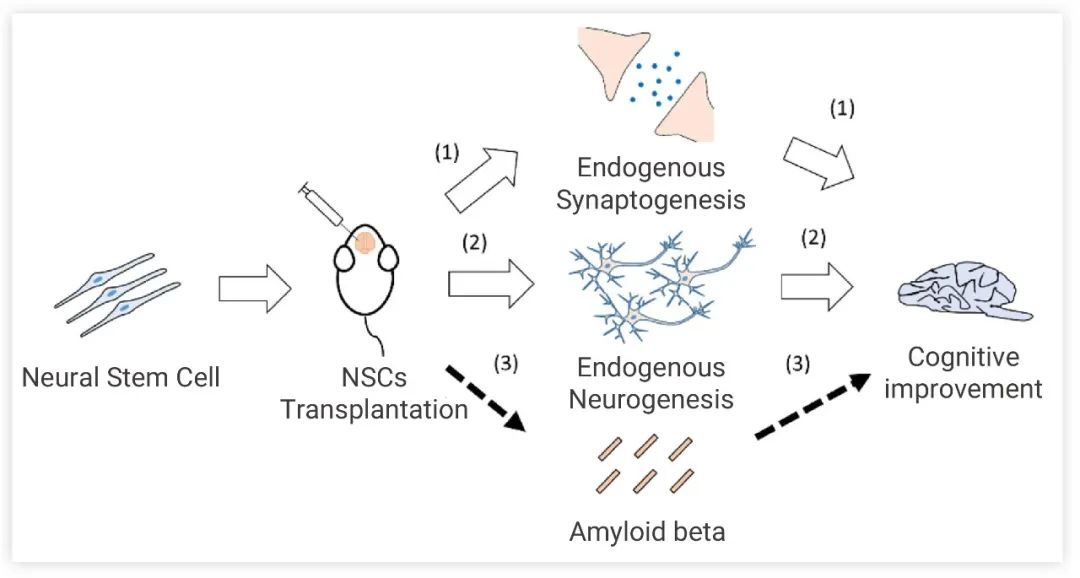 圖 4. NSCs 移植途徑及認知障礙修復機制:觸發內源性突觸形成、內源性神經形成影響行為表現、調節 Aβ 聚集[6]
圖 4. NSCs 移植途徑及認知障礙修復機制:觸發內源性突觸形成、內源性神經形成影響行為表現、調節 Aβ 聚集[6]
接下來小 M 來介紹一個在 APPswe/PS1dE9 雙轉基因小鼠中移植 NSCs 治療 AD 的例子~
蘇州納米所王強斌團隊開發了一種能夠穩定和持續表達腦啡肽酶 (NEP) 的基因工程 NSCs。NSCs 經改造后,在細胞膜上表達的 NEP 和釋放的胞外載體可持續降解 Aβ,提高 NSCs 和內源性神經細胞的存活率。PPAR-siSOX9 納米制劑可通過協同調節 Wnt/β-catenin 和 RA 信號通路來提高 NEP-NSC 的分化效率。
研究者們將改造的 PPAR-siSOX9 納米制劑-NEP-NSCs 立體定向移植到 APPswe/PS1dE9 雙轉基因 AD 小鼠模型的海馬體中(海馬體是大腦中最容易受到 AD 病理學損傷形成的區域)。與對照組相比,經過治療后的 AD 小鼠具有更高活力和更高密度的神經元,并且具有更高的存活率。治療一個月后,大腦中的病理性 Aβ 聚集的斑塊面積和大小也顯著降低 (圖 5)。在 Morris 水迷宮實驗中證明,納米制劑-NEP-NSCs 治療小鼠更頻繁地穿過平臺并在目標象限中游泳的時間更長。這些結果表明,經改造后的納米制劑-NEP-NSCs 在 AD 小鼠模型中能更有效地降解 Aβ 以及再生神經元,逆轉了小鼠記憶與學習缺陷。
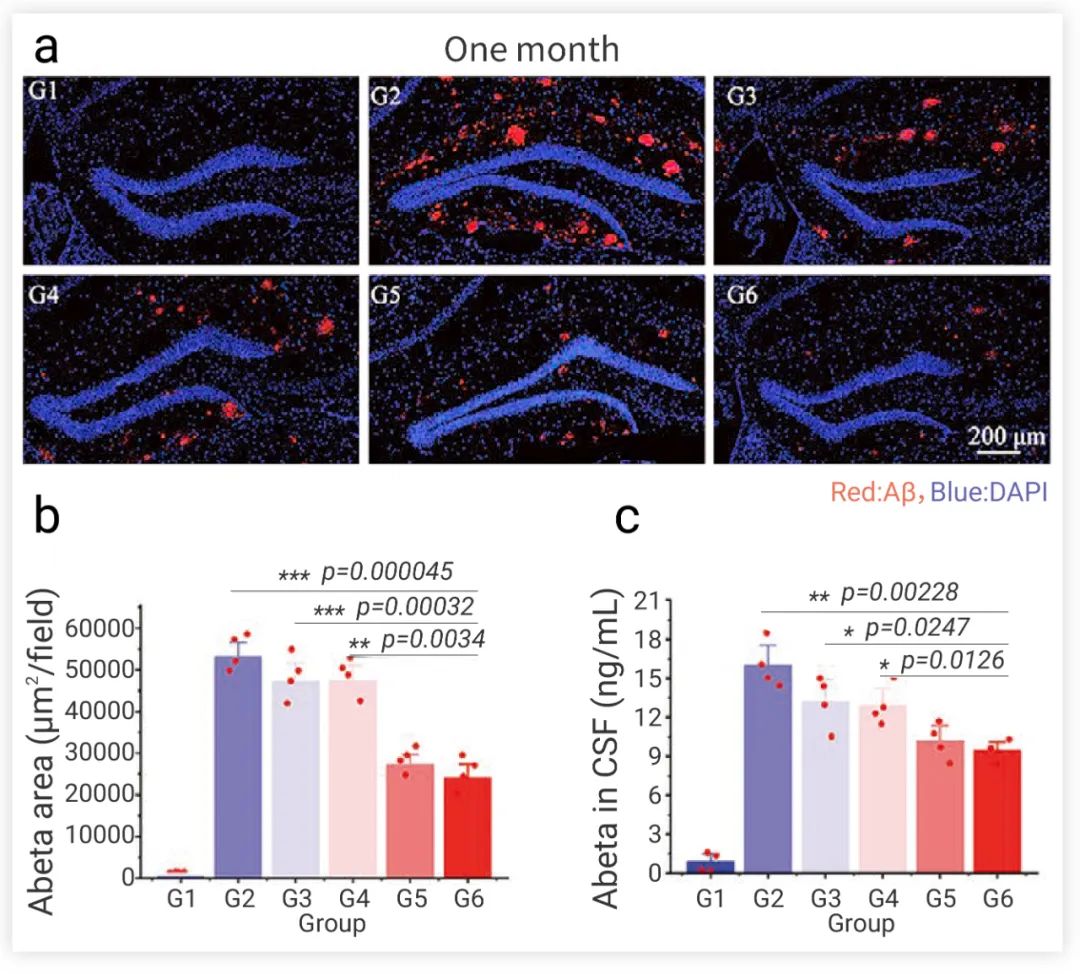
圖 5. NSCs 移植治療后 AD 小鼠大腦中 Aβ 定量檢測。G1:健康小鼠;G2:PBS 處理組;G3:NSC 治療組;G4:納米制劑-NSC 治療組;G5:NEP–NSC 治療組;G6:納米制劑-NEP-NSC 治療組[7]
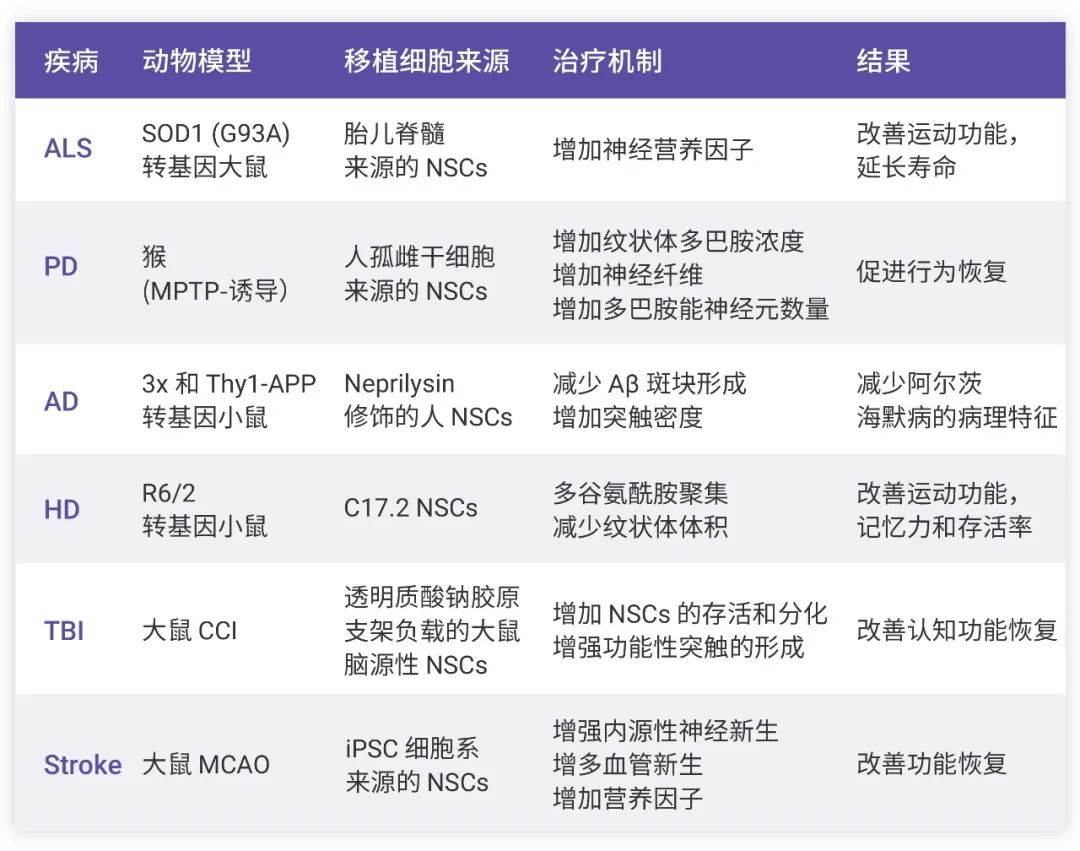
除了阿爾茨海默癥,在其他疾病如帕金森病 (PD)、肌萎縮側索硬化癥 (ALS)、亨廷頓病 (HD)、中風、創傷性腦損傷 (TBI) 等神經系統疾病的治療中,NSCs 移植也表現出巨大的潛力。
17038899, 27213850, 25022790, 18683244, 27562609, 25132189
|
相關產品 |
|
一種有效的有絲分裂因子,通過與細胞表面受體——表皮生長因子受體 (EGFR/ErbB) 的高親和力結合發揮作用。EGF 在眾多類型細胞的生長、增殖和分化中發揮著重要作用,常用于 NSCs 的增殖培養。 |
|
一種內皮細胞和成纖維細胞的有效有絲分裂原,有廣泛的生物學功能,包括有絲分裂發生、細胞存活、轉移形成,常用于 NSCs 的培養,bFGF 誘導神經干細胞增殖。 |
|
NGF 是神經營養因子中發現最早的因子,迄今為止研究最徹底,具有提供神經元營養和促進突觸生長的雙重生物學功能。 |
|
一種神經營養因子,能夠與 TrkB 受體結合,調節神經發育過程,包括神經元的存活,分化,及突觸可塑性。BDNF 能刺激 NSCs 增殖,顯著增加 NSCs 向神經元和少突膠質細胞的分化。 |
|
神經營養因子家族中的一員,能夠與兩種不同受體結合,高親和性受體 TrkB 和低親和性受體 p75NTR。 |
|
在神經系統中廣泛表達。NT-3 能促進 BM-NSCs 增殖和分化為膽堿能神經元,并增加上清液中乙酰膽堿 (ACh) 的水平。 |
|
一種多能性神經營養因子,高濃度的CNTF通過 JAK-STAT 信號通路作用,將細胞分化為星形膠質細胞。低濃度 CNTF 作為另一種神經元誘導劑。這種誘導劑屬于造血細胞因子超家族,通過與 CNTF 受體 α (CNTFRα) 和 gp130 結合發揮作用,最終募集白血病抑制因子受體 β (LIFRβ) 并激活絲裂原活化蛋白激酶通路。 |
|
一種有效的促有絲分裂因子 (有絲分裂原),能調節結締組織和發育中神經系統的細胞生長和分裂。甲狀腺激素 (T3) 和血小板衍生生長因子 (PDGF) 的組合通過 Wnt/β 促進少突膠質細胞的分化-連環蛋白信號通路。 |
|
Hedgehog 家族分泌信號蛋白的一員,在成體干細胞增殖中起作用。SHH 是細胞增殖、神經發生和神經元成熟的正向調節劑[17]。 |
|
IGF-I 在大腦發育過程中表達,被認為作用于所有神經細胞。IGF-I 是調節 NSCs 活化的關鍵。 |
|
誘導神經前體細胞 (iNPCs) 衍生多巴胺能神經元 (DA) 生成的關鍵因子。 |
|
一種神經營養因子,能促進中腦多巴胺能神經元的存活分化,支持人多能干細胞來源的神經祖細胞向神經元分化。 |
|
用于染色細胞核。 |
參考文獻
1. Yuewen Tang, L Cheng, et al. Current progress in the derivation and therapeutic application of neural stem cells. Cell Death Dis. 2017 Oct 12;8(10):e3108.
2. Yanjing Zhu, Ruiqi Huang, Liming Cheng, Rongrong Zhu, et al. Deep learning-based predictive identification of neural stem cell differentiation.Nat Commun. 2021 May 10;12(1):2614.
3. Roberta De Gioia, Stefania Corti, et al. Neural Stem Cell Transplantation for Neurodegenerative Diseases. Int J Mol Sci. 2020 Apr 28;21(9):3103.
4. Yuan Zhang, Xin-Yu Lu, Giacomo Casella, Jing Tian, Xing Li, et al. Generation of Oligodendrocyte Progenitor Cells From Mouse Bone Marrow Cells.Front Cell Neurosci. 2019 Jun 5;13:247.
5. Kyung-Chul Choi,Do-Sung Yoo, et al. Effect of Single Growth Factor and Growth Factor Combinations on Differentiation of Neural Stem Cells. J Korean Neurosurg Soc. 2008 Dec; 44(6): 375-381.
6. Hayashi Y, Tsai KJ, et al. Effects of neural stem cell transplantation in Alzheimer's disease models. J Biomed Sci. 2020 Jan 27;27(1):29.
7. Huang D, Wang Q, et al. A Nanoformulation-Mediated Multifunctional Stem Cell Therapy with Improved Beta-Amyloid Clearance and Neural Regeneration for Alzheimer's Disease. Adv Mater. 2021 Apr;33(13):e2006357.



