
ŌŌõŖģ¯ĐŪīĩ°°×ŊMŊŌĘžĖĮģųŪDŌÆøŌŌõŖģ¯ÔÚ×ĐÎæĮōžúÉúÎīĤĩÄ×÷ĶÃ

°ląíÆÚŋ¯ŖēEnviron Microbiol Rep
°ląírégŖē2021Äę4ÔÂ13ČÕ
Ö÷ŌĒŧŧĐgŖēŌŌõŖģ¯ĐŪīĩ°°×ŊMW
ĶąžîĐÍŖēŧžú
| ÕĒŌĒ
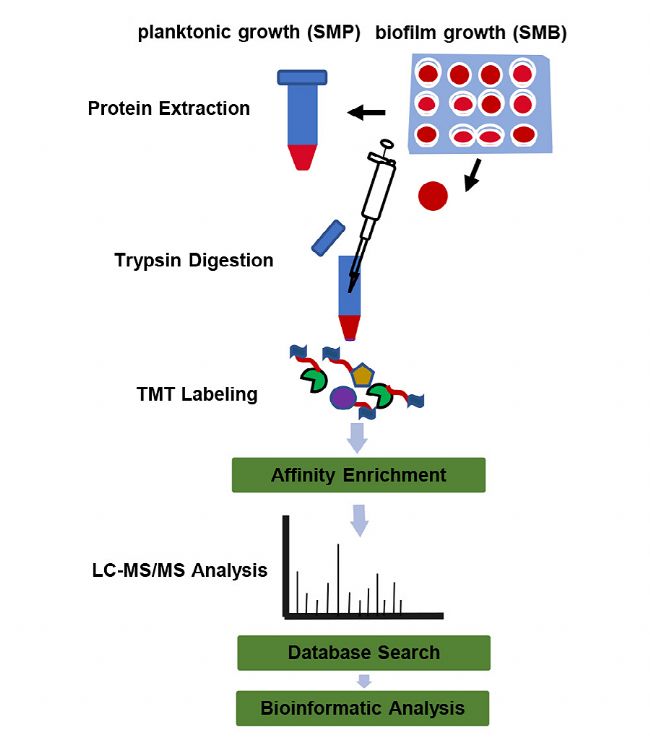
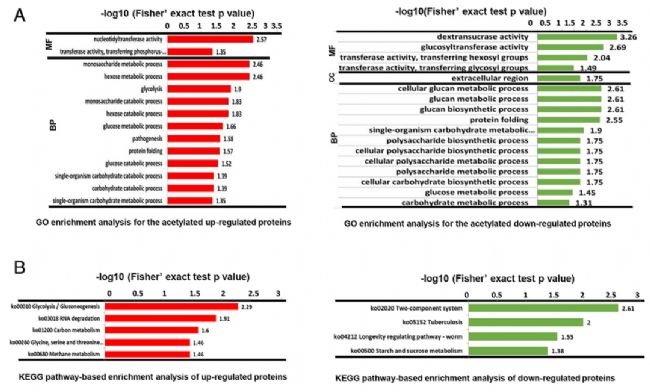
×ĐÎæĮōžúÄÜĶĐЧĩØĀûĶÃĘŗÎīÖĐĩÄÕáĖĮŽaÉú°ûÍâļāĖĮŖŦļø°ûÍâļāĖĮÖ÷ŌĒĘĮͨß^ĖĮģųŪDŌÆø(Gtfs)ĩÄ×÷ĶÃēĪŗÉĩÄĄŖąžŅĐžŋĻ×ĐÎæĮōžúĩÄŌŌõŖģ¯ĐŪīĩ°°×ßMĐĐÁËčbļ¨ŖŦ˛ĸĻ×ĐÎæĮōžúÉúÎīĤÉúéL(SMB)ē͸ĄĶÎÉúÎīĤÉúéL(SMP)ĩÄŌŌõŖģ¯ĐŪīĩ°°×ßMĐĐÁËļ¨ÁŋˇÖÎöŖŦ˛ÉĶÃTMTËĶēÍKacĶHē͸ģŧ¯ŊYēĪ¸ßˇÖąæÂĘŲ|×Vļ¨Áŋĩ°°×Ų|ŊMWĩġŊˇ¨ļ¨Áŋĩ°°×Ų|ŌŌõŖģ¯ĩÄĶB×ģ¯ŖŦš˛čbļ¨ÁË445ĩ°°×ÖĐĩÄ973ŌŌõŖģ¯ÎģücŖŦÆäÖĐļ¨ÁŋÁË302ĩ°°×ÖĐĩÄ617ŌŌõŖģ¯ÎģücĄŖ
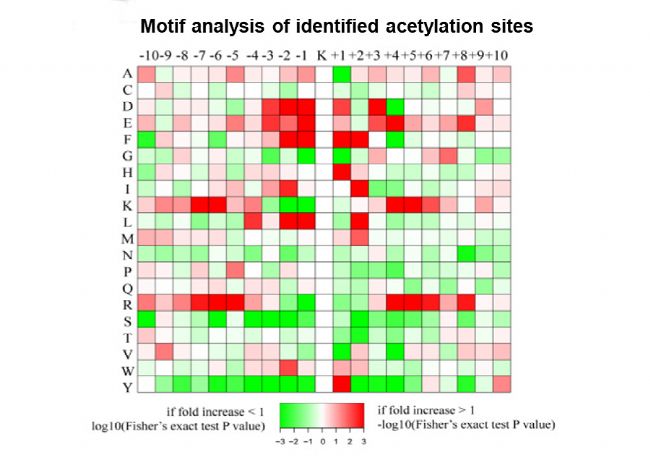
ŋķwˇÖÎöąíÃ÷ŖŦ22.7%ĩÄĩ°°×Ų|°lÉúÁËŌŌõŖģ¯ĄŖÔÚSMBĩÄļ¨Áŋĩ°°×ÖĐŖŦĶĐ56ÎģücŲ°ąËáĩÄŌŌõŖģ¯ŗĖļČÉũ¸ßŖŦ52ÎģücŲ°ąËáĩÄŌŌõŖģ¯ŗĖļČŊĩĩÍĄŖÔÚ×ĐÎæĮōžúĩÄŌŌõŖģųŊMÖĐŖŦčbļ¨ĩÃĩŊ6ī@Öø¸ģŧ¯ĩÄģųÔĒŖŦˇÖeéKac****KĄĸKacFĄĸKac****RĄĸKacYĄĸKacHĄĸF*KacĄŖ´ËÍâŖŦģųĶÚKEGGÍžŊĩĸģŧ¯ˇÖÎöąíÃ÷ŖŦĖĮŊÍŊâ/ĖĮŽÉúēÍRNAŊĩŊâī@Öø¸ģŧ¯ĄŖĖØeĘĮÔÚ×ĐÎæĮōžúÉúÎīĤÖĐŖŦŌŌõŖģ¯Ų°ąËáĩ°°×ĪÂÕ{×îļāĩÄĘĮÆĪĖĮģųŪDŌÆø- SIĄĸÆĪĖĮģųŪDŌÆø- IēÍÆĪĖĮģųŪDŌÆø- SŖŦß@ŋÉÄÜŊŌĘžÁËÉúÎīĤ°lĶũß^ŗĖÖĐÆĪĖĮģųŪDŌÆøšĻÄÜÕ{ŋØĩÄęPé]CÖÆĄŖ
| ŅĐžŋŊYšû
1.ĻÍģ×æĮōžúÖĐËųĶĐŌŅčbļ¨ĩÄŲ°ąËáŌŌõŖģ¯ĩ°°×ßMĐĐžCēĪˇÖÎö
ͨß^LC-MS/MSzy×ĐÎæĮōžúÖĐ°lÉúŌŌõŖģ¯ĐŪīĩÄĩ°°×ŖŦš˛čbļ¨ŗö445ĩ°°×ÖĐ973ŌŌõŖģ¯ÎģücŖŦÆäÖĐ306ĩ°°×ÖĐ617ŌŌõŖģ¯ÎģücßMĐĐÁËļ¨ÁŋˇÖÎöĄŖÆäÖĐĶĐ56ÎģücŲ°ąËáĩÄŌŌõŖģ¯ŗĖļČÉũ¸ßŖŦ52ÎģücŲ°ąËáĩÄŌŌõŖģ¯ŗĖļČŊĩĩÍĄŖ
2. ×ĐÎæĮōžúŌŌõŖģ¯ĩ°°×ĩÄŧ°ûļ¨ÎģēÍšĻÄܡֲŧ
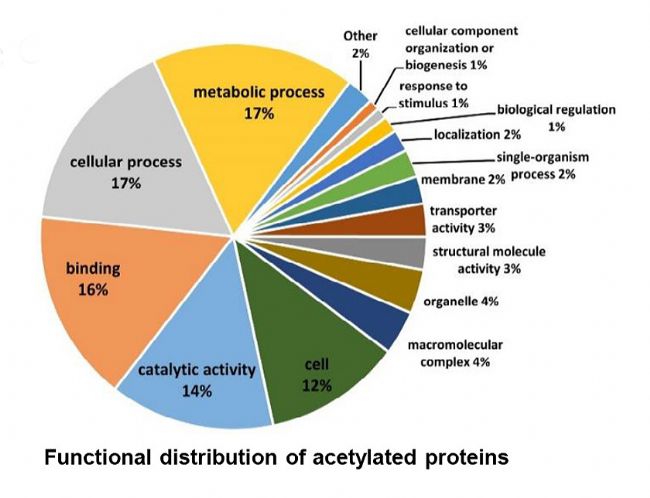
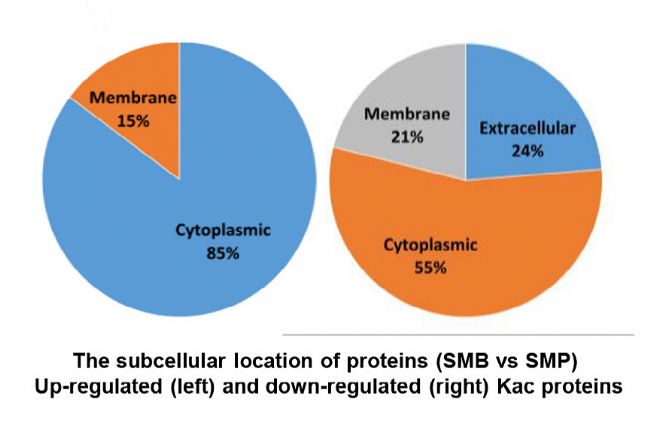
×ĐÎæĮōžúē͸ĄĶÎĪāąČŖŦĻÆä˛îŽĩÄŌŌõŖģ¯ĩ°°×ßMĐĐŧ°ûļ¨ÎģēÍšĻÄܡÖÎöŖŦÆäÖĐ×ĐÎæĮōžú85%ĩÄŌŌõŖģ¯ĩ°°×ˇÖ˛ŧÔÚŧ°ûŲ|ÖĐŖŦ15%ˇÖ˛ŧÔÚĤÖĐŖŦļø¸ĄĶÎÉúÎīÖĐŌŌõŖģ¯ĩ°°×55%ˇÖ˛ŧÔÚŧ°ûŲ|ÖĐŖŦ24%ˇÖ˛ŧÔÚŧ°ûÍâŋÕégŖŦ21%ˇÖ˛ŧÔÚĤÖĐĄŖ¸ųūšĻÄܡֲŧŖŦ×ĐÎæĮōžúÖĐ17%ĩÄŌŌõŖģ¯ĩ°°×
ĸÅc´úÖxß^ŗĖŖŦ17%
ĸÅcŧ°ûß^ŗĖŖŦ17%
ĸÅcŊYēĪŖŦ14%
ĸÅc´ßģ¯ģîĐÔŖŦ12%
ĸÅcŧ°ûĄŖÉĪĘöŊYšûąíÃ÷ŖŦŲ°ąËáŌŌõŖģ¯ĩ°°×ÔÚ×ĐÎæĮōžúÖĐVˇēˇÖ˛ŧŖŦĖáĘžŌŌõŖģ¯ĐŪīŋÉÄÜÔÚÉúÎīĤĐÎŗÉÖĐÆđÖØŌĒĩÄÕ{ŋØ×÷ĶÃĄŖ
3. ÔÚ¸ĄĶÎēÍ×ĐÎæĮōžúÉúÎīĤŧ°ûÖвîŽŌŌõŖģ¯ĩ°°×ĩÄšĻÄܸģŧ¯
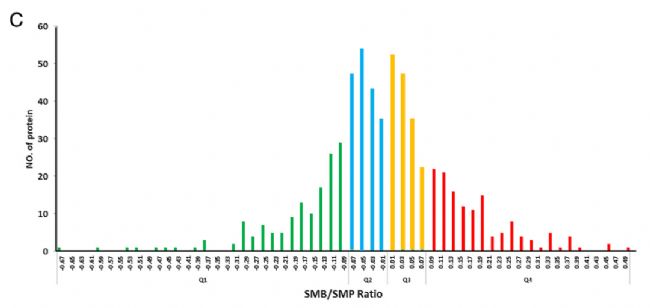
éÁËŅĐžŋŌŌõŖģ¯ĐŪīĘĮˇņ¸Ä×ĩ°°×ĩÄšĻÄÜŖŦßMĐĐÁËģųĶÚGOĩĸģŧ¯ˇÖÎöĄŖÔÚĖŧËŽģ¯ēĪÎī´úÖxē͡ÖŊâ´úÖxŖŦŌÔŧ°ÆĪžÛĖĮ´úÖxß^ŗĖžųī@Öø¸ģŧ¯ÔÚÕ{ŋØĩ°°×ÖĐĄŖģųĶÚKEGGͨ¡ĻÕ{ŋØĩ°°×(SMB vs SMP)ĩĸģŧ¯ˇÖÎöąíÃ÷ŖŦī@Öøĩĸģŧ¯ĘĮĖĮŊÍŊâ/ĖĮŽÉúēÍRNAŊĩŊâĄŖĸ¸÷ŌŌõŖģ¯ëÄĩÄSMB/SMP TMTąČÖĩŪDģ¯éz-scoreŖŦ¸ųū25%Ąĸ50%Ąĸ75%Čũ˛ģÍŦ°ŲˇÖÎģĩÄÕũBˇÖ˛ŧĀÛˇeÃÜļČē¯ĩÔOÖÃz-scoreĩÄŊØÖšÖĩĄŖļ¨ÁŋŊYšûī@ĘžŖŦ´ķ˛ŋˇÖŌŌõŖģ¯ĩ°°×ˇÖ˛ŧÔÚQ2(25 50%)ēÍQ3(50 75%)ˇÖÎģĩÖĐĄŖ
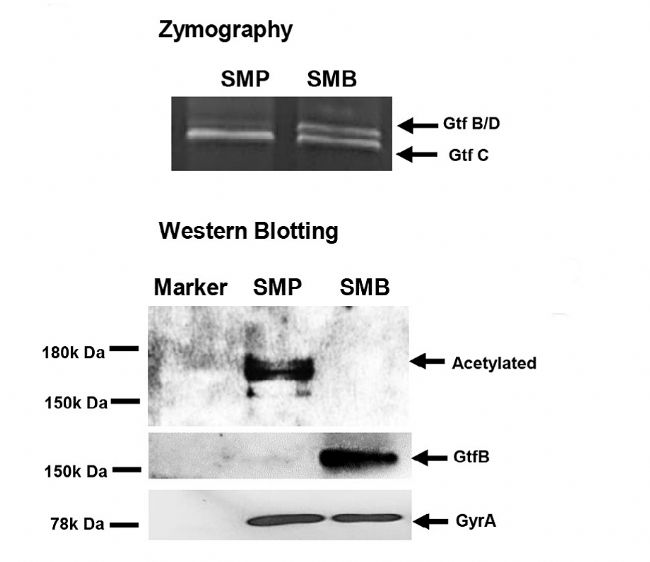
4. WB ō×C
Ļ˛îŽŌŌõŖģ¯ĩ°°×ĩÄļ¨ÁŋˇÖÎöąíÃ÷ŖŦÔÚ×ĐÎæĮōžúÉúÎīĤÖĐŖŦŌŌõŖģ¯Ų°ąËáĩ°°×ĪÂÕ{×îļāĩÄĘĮÆĪĖŅĖĮģųŪDŌÆø-SIĄĸÆĪĖŅĖĮģųŪDŌÆø- IēÍÆĪĖŅĖĮģųŪDŌÆø-SĄŖͨß^ WB ō×CŌŌõŖģ¯ĐŪīËŽÆŊŖŦÅc¸ĄĶÎÉúÎīÉúéLĪāąČŖŦ×ĐÎæĮōžúÉúÎīĤÖĐĩÄŌŌõŖģ¯ÆĪĖŅĖĮģųŪDŌÆøī@ÖøĪÂÕ{ĄŖ
| ŅĐžŋŊYÕ
ÔÚ׎æĮōžúÖĐčbļ¨ÁË445ĩ°°×ÖĐ973ŌŌõŖģ¯ÎģücĄŖÆäÖĐ22.7%ĩÄĩ°°×Ų|°lÉúÁËŌŌõŖģ¯ŖŦÖ÷ŌĒ°lÉúÔÚ6ī@Öø¸ģŧ¯ĩÄģųÔĒÉĪŖŦČįKac****KĄĸKacFĄĸKac****RĄĸKacYĄĸKacHēÍF*KacĄŖ
ŋĩÄíÕfŖŦÔÚÉúÎīĤëAļÎŌŌõŖģ¯ĩ°°×ĩÄĩÁŋÔöŧĶĄŖļøŌŌõŖģ¯/ČĨŌŌõŖģ¯¸Ä×ĩÄĩ°°×Ų|Ö÷ŌĒ
ĸÅcĖĮ´úÖxē͡ÖŊâ´úÖxŖŦŌÔŧ°ÆĪžÛĖĮĩÄ´úÖxß^ŗĖĄŖĖØeĘĮ×ĐÎæĮōžúÉúÎīĤÖĐŌŌõŖģ¯ËŽÆŊĪÂÕ{×îļāĩÄĩ°°×ŖŦČįÆĪĖŅĖĮģųŪDŌÆø- SIĄĸÆĪĖŅĖĮģųŪDŌÆø- IēÍÆĪĖŅĖĮģųŪDŌÆø- SŖŦŋÉÄÜ´ŲßMÉúÎīĤĩÄĐÎŗÉĄŖß@ĐŠĖĮģųŪDŌÆøŋÉÄÜĘĮũxũXÖίĩÄÔÚ°ĐücĄŖ
ÎÄĢIÔÎÄĪÂŨdæŊĶ
https://pan.baidu.com/s/1qhz90m7Y-6Pt0LzvdaowyQ
ĖáČĄ´aŖēksv9

- ŌíēÍÉúÎīSNPģųŌōˇÖĐÍŧŧĐgŖēŊŌÃØģųŌōÃÜ´aŖŦĖŊ¤ŊĄŋĩ֎¡
- ´ŲÄIÉĪĪŲƤŲ|ŧ¤ËØáˇÅŧ¤ËØŖ¨CRHŖŠÔÚĖŊË÷ˇĘÅÖCÖÆÖĐĩÄŅĐžŋ
- ÃÚÄōĪĩŊyŧ˛˛ĄĩÄęPæIÖί°ĐücÖŽV2RĩÄŊéŊBŧ°ĪāęPō
- ŧ¤šâš˛žÛŊšī@ÎĸįRÖúÁĻÄ[ÁöŌÖÖÆŠŅĐžŋ
- ŋÕégļāŊMWŊâ´aÄ[ÁöÎĸhžŗŖŦÖúÁĻŧĶËŲžĢĘÖίŅĐžŋ
- ĀûĶÃÎŧ°ûŪDäŊMŧŧĐgŊâÎöļāˇNÃâŌßŧ°ûĻ´ķÁŋŧ°ûŌō×ĶĩÄíĒ
- EmulateÆ÷šŲĐžÆŦÖúÁĻŅĐžŋËÎīŌũÆđ¸ÎpûĩÄCÖÆ
- šĮŲ|ĘčËÉēÍŧ× îÅÔĪŲšĻÄÜpÍËęPæI°ĐücPTH1RĩÄŊYšĻÄÜŧ°ĪāęPōŊéŊB
- ÉúÎīĐžÆŦÍÆŗö"ĐžŋÕŌģĖ"ˇŊ°¸ÖØËÜŋÕégļāŊMWĐ¡ļĘŊ
- IPHASEÕnĖÃé_ÕnĀ˛ŖēŧžúģØÍÍģ×ÔōŗŖŌî}ÅcŊâ´đ
- ÉúÎīĐžÆŦÅcĐžŗŦÉúÎīÂēĪÍÆŗö"ĐžŋÕŌģĖ"ļāŊMWˇŊ°¸
- ÉúÎīátËzyÅcÔŠšŠĒÉĖŌíēÍÉúÎīĪōČĢøÕĐÄŧŊäNÉĖ
- žÉĪÕnŗĖŖēØiÄcĩĀšÚ ļžÅcČËķwoÂĩÄ´úÖxļˇ
- Ōģ×÷Öą˛ĨŖēŋšĐÔĩíˇÛͨß^ÖØËÜÄcĩĀÎĸÉúÎīČēžŊâˇĘÅÖ
- SBC&ÄŖģųÉúÎī2024ÄęĩÚŌģÆÚîÆ÷šŲŗõŧÅāĶ°āŅûÕē¯
- SBCŅûÄú ĸŧĶ2024ŋÕégļāžSŊMWĐÂŪDģ¯ŅĐĶū


