
外泌體的介紹和檢測提取機制

外泌體 (Exosome) 是什么?
所有原核和真核細胞在生理正常或異常時,均可通過出芽的方式釋放各種各樣的膜包裹的囊泡 (Extracellular vesicles, EVs) 到胞外環境中,如外泌體、微囊泡、凋亡小體。胞外囊泡可廣泛的分為兩類,核外顆粒體 (Ectosome) 和外泌體。核外顆粒體是從質膜脫離的外出芽小泡,包括直徑在 ~50 nm-1 μm 的微泡、微粒和大囊泡。
外泌體是細胞核內體途徑起源的、直徑在 ~40-160 nm (平均 100 nm) 的囊泡。外泌體的產生涉及到一個獨特的細胞內調節過程,這可能決定了它們的組成,從而決定了它們的功能。
外泌體的產生和分泌
外泌體的產生涉及質膜的雙重內陷、和含有腔內小泡 (Intraluminal vesicles, ILVs)、細胞內多囊泡體 (Multivesicular bodies, MVBs) 的形成。ILVs 通過 MVBs 與質膜的融合和胞吐,最終以外泌體的形式分泌到胞外。
質膜的第一次內陷形成一個杯狀結構,其中包含細胞表面的蛋白與胞外一些可溶性蛋白,這個過程形成了早期內涵體 (Early-sorting endosome, ESE)。ESEs 可發展為成熟的晚期內涵體 (Late-sorting endosomes, LSEs),并最終生成 MVBs。MVBs 是通過質膜雙凹形成的,這一過程導致 MVBs 含有多個 ILVs (未來的外泌體)。MVBs 可以與溶酶體或自噬體融合被降解,或與質膜融合以釋放所含的 ILVs 為外泌體。
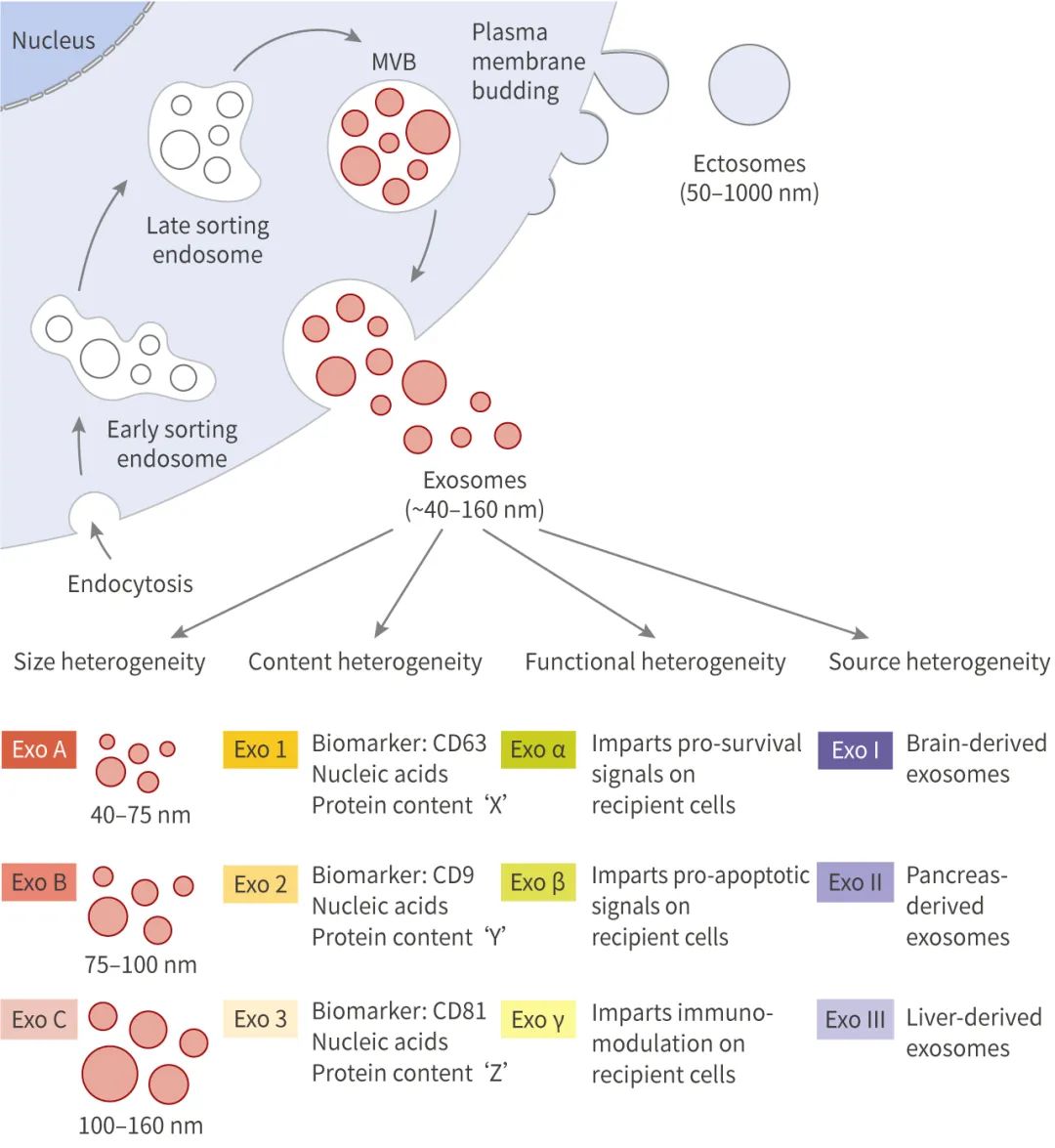
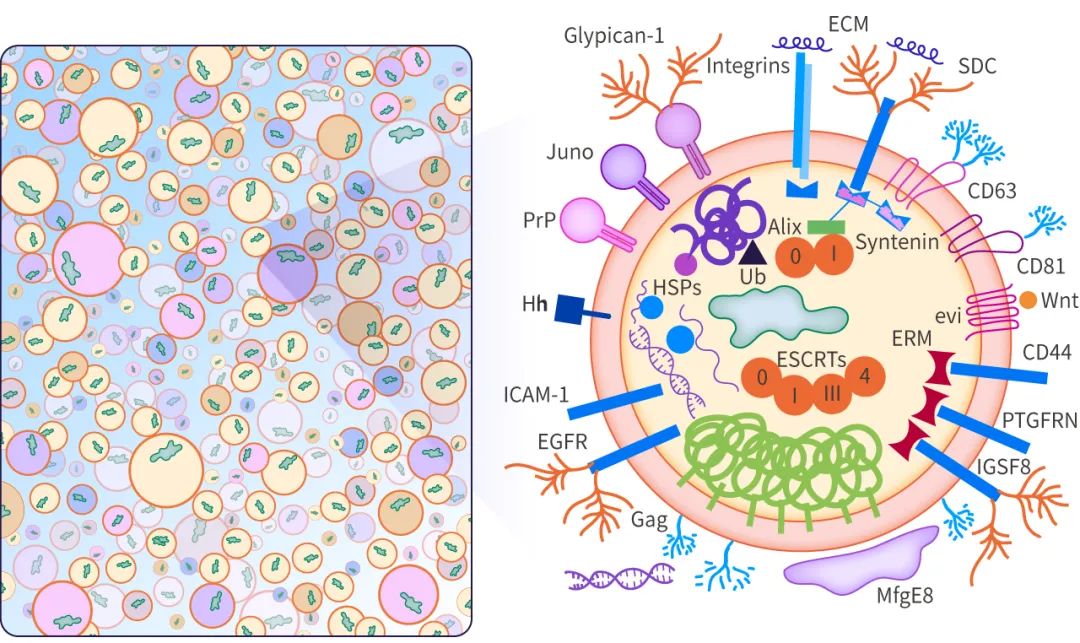
外泌體的起源和形成方式導致其內容物、形狀、大小均有差異,從而會影響受體細胞的不同功能,這種特性被稱為外泌體的異質性。外泌體可通過其大小和表面特異性標記被識別,包括 TSG101、Alix、Flotillin-1、CD63、CD9 等。
外泌體分泌途徑的分子機制
■ ESCRT
說起分泌和轉運,不得不提轉運所需的內涵體分選復合體 (Endosomal sorting complex required for transport, ESCRT)。ESCRT 是一個蛋白家族,在 MVBs 膜上連續結合復合物 (ESCRT-0, -I, -II 和 -III),其中 ESCRT-0 通過泛素化依賴途徑調控內容物聚集,ESCRT-I 和 ESCRT-II 誘導芽的形成,ESCRT-III 驅動小泡的脫落,而輔助蛋白 (特別是 VPS4 ATPase) 允許 ESCRT 系統的解離和循環。
ESCRT-0:ESCRT-0 復合體 (HRS 和 STAM) 不參與出芽和膜分離的過程,因此可作為追蹤外泌體來源的依據。HRS (Hepatocyte growth factor-associated tyrosine kinase) 識別單泛素化的蛋白,并與泛素化部分結合將蛋白靶向到 ILVs 上 (蛋白分選到 ILVs 中的過程主要取決于泛素化),并與 STAM、Eps15 和 Clathrin 形成復合體。隨后 HRS 招募腫瘤易感基因 TSG101 (ESCRT-I 的組分),ESCRT-0 和 ESCRT-I 一起負責泛素化蛋白的分選。
ESCRT-II :ESCRT-II 是一個異四聚體的蛋白復合物,由 Vps36、Vps22 和雙拷貝的 Vps25 組成。Vps36 的 N 末端可與泛素 (內容物蛋白或其它 ESCRT 上蛋白)、以及 ESCRT-I 的 C 末端結合。ESCRT-I/II 的超復合體被證明在 ILVs 的膜變形中發揮重要作用。然后,ESCRT-II 的 Vps25 亞基和 ESCRT-III 的 Vps20 亞基結合,啟動 ESCRT-III 工作。
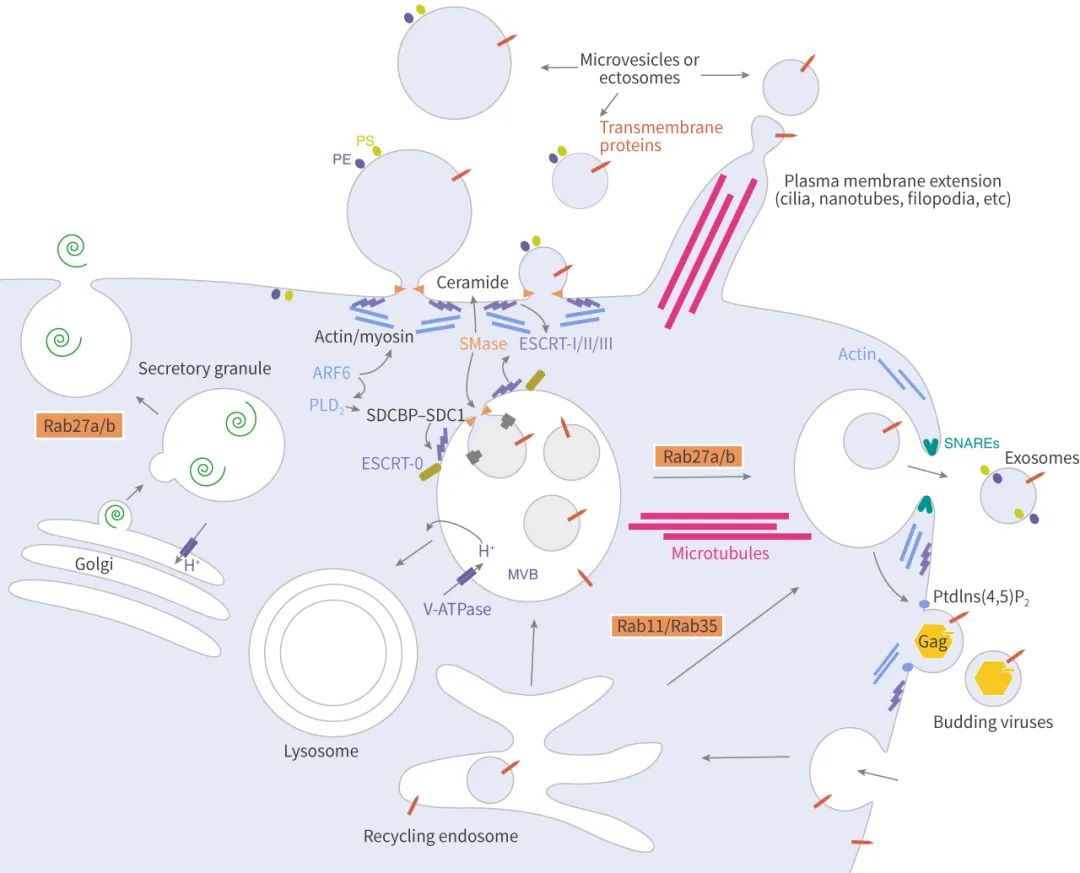
ESCRT-III:以酵母為例,ESCRT-III 由 Vps20、Snf7 (Vps32)、Vps24 和 Vps2 四個核心亞基、以及輔助蛋白 Did2、Vps60、和 Ist1 組成,在細胞質中,這些亞基都是不活躍的單體,一旦結合膜上,它們形成一個瞬態的 ESCRT-III 異聚合物。前面說到 ESCRT-II 的 Vps25 亞基和 ESCRT-III 的 Vps20 亞基結合, 然后 Snf7 被 Vps24 和 Vps2 包裹 (像帽子蓋住),與 Vps20 形成復合體。ESCRT-III 促進膜分離,促使囊泡斷離。
■ Rab GTPases
Rab 家族的小 GTPases 成員在細胞內 ILVs 的轉移中發揮了明確的作用,也參與了 MVBs 與質膜的對接、以及外泌體的釋放。其中 Rab27a 和 Rab27b 作用于 MVBs (也叫 MVE,Multivesicular endosomes) 與質膜的對接。Rab27a 抑制后,MVBs 的大小顯著增加,而抑制 Rab27b 后導致 MVBs 的向核周圍區域的重新分布,此外,Rab27 兩個效應因子 Slp4 (SYTL4) 和 Slac2b (EXPH5) 被抑制后同樣抑制外泌體的分泌且分別導致 Rab27a 和 Rab27b 的沉默。
■ SNAREs
研究外泌體生物發生的一個主要目標是識別 MVBs 與質膜融合所需要的 SNARE (可溶性 N-乙基馬來酰亞胺敏感融合連接蛋白受體) 復合物。YKT6 SNARE 被證明是分泌含 Wnt 的外泌體所必需的,而 Syx-5 (線蟲同源物 Syntaxin-5),通過 Ral-1 小 GTPase 使 MVBs 靶向質膜。另外,Syntaxin-1A 是神經元特異性的 SNARE,參與突觸囊泡的分泌,能影響果蠅的外泌體分泌。
■ 其它
ARF6 是 ILVs 出芽和外泌體生物合成的調節因子,該調節作用依賴于 ARF6 的效應器 PLD2。
Doa4 N 末端具有一個相當于 MIT 結構域的 α 螺旋結構,直接與 ESCRT-III 的 Vsp20 亞基中類似 MIM1 的序列結合,可促進 ILVs 的形成。
外泌體被生產出來后去哪里了呢?小編告訴你,它們可能被別的細胞“吃了”!
外泌體的攝取
目前還不清楚外泌體是否必須被免疫細胞和非免疫細胞內化以引起細胞反應。根據現有研究,外泌體被攝取內化的途徑主要有以下幾種。
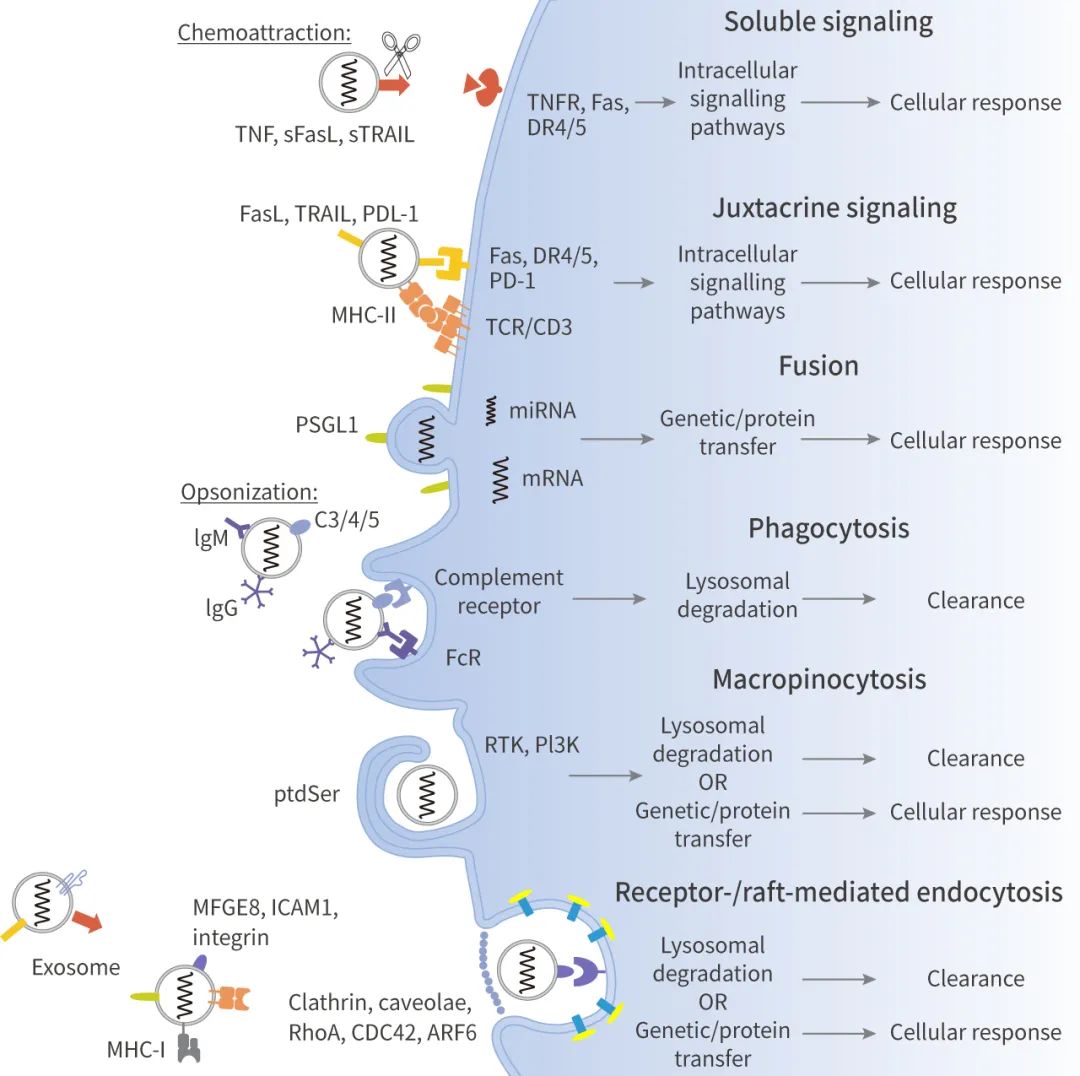
◎ 可溶性信號通路:對外泌體表面的配體進行蛋白水解切割或選擇性剪接。比如:膜結合的 FasL、TRAIL 和 TNF 可以被金屬蛋白酶切割,形成可溶性細胞因子。
◎ 近分泌信號通路:外泌體和靶細胞表面的配體和受體鄰近,通過信號傳導途徑發揮作用。比如:培養的胎盤外植體或孕婦血漿中的外泌體膜上具有 FasL 和 TRAIL,并通過 NF-κB、CD3ζ 和 JAK3 的下調,來誘導 Jurkat T 細胞凋亡。
◎ 融合:例如,研究證實單核細胞來源的微泡可以與活化的血小板的質膜融合,并將蛋白轉移到受體細胞,如組織因子和 P-選擇素糖蛋白配體-1 (PSGL-1)。
◎ 吞噬作用:吞噬是一種肌動蛋白介導的機制,需要特定的調理素蛋白受體 (如 FcR 和補體受體)、清道夫受體或 Toll 樣受體的參與。可由巨噬細胞和樹突狀細胞這類“專業細胞”完成,也可由 γδT 等“非專業”細胞完成。例如,單核/巨噬細胞能夠更有效地吞噬內化來自紅血球 (K562) 和 T 細胞白血病 (MT4) 細胞的外泌體。外泌體的吞噬作用依賴于肌動蛋白細胞骨架、磷脂酰肌醇 3 激酶 (PI3K) 和 Dynamin2。
◎ 巨胞飲:少突細胞來源的外泌體表面的磷脂酰絲氨酸 (PtdSer) 激活了不具備抗原呈遞能力的小膠質細胞/巨噬細胞亞群的大胞吞。外泌體的巨胞飲作用依賴于 Na+ 和 PI3K,EIPA 和 LY294002 分別抑制 Na+-H+ 離子交換和 PI3K 活性,減少外泌體攝取。
◎ 受體和脂筏介導的內吞:這兩個途徑需要外泌體表面的配體與質膜上的特定受體結合,在細胞質膜上分別需要存在富含膽固醇和鞘脂的微區。前者也稱為網格蛋白介導的內吞作用。后者包括小窩介導的內吞作用,以及 RhoA-、CDC42-和 ARF6-調控的、獨立于網格蛋白和小窩的內吞作用,它們利用 Dynamin、Flotillin 和/或 Rab 蛋白的不同組合。如大鼠腎上腺髓質 (PC12) 腫瘤細胞釋放的外泌體可通過網格蛋白介導的內吞作用部分內化。
參考文獻
↓ 下滑查看更多文獻
1. Raghu Kalluri, et al. The biology , function , and biomedical applications of exosomes. Science. 2020 Feb 7;367(6478):eaau6977.2. Mathilde Mathieu, et al. Specificities of secretion and uptake of exosomes and other extracellular vesicles for cell-to-cell communication. Nat Cell Biol. 2019 Jan;21(1):9-17.
3. Suresh Mathivanan, et al. Exosomes: extracellular organelles important in intercellular communication. J Proteomics. 2010 Sep 10;73(10):1907-20.
4. Lucio Barile, et al. Exosomes: Therapy delivery tools and biomarkers of diseases. Pharmacol Ther. 2017 Jun;174:63-78.
5. Kelly J. McKelvey, et al. Exosomes: Mechanisms of Uptake. J Circ Biomark. 2015 Jan-Dec; 4: 7.
6. Marina Colombo, et al. Analysis of ESCRT functions in exosome biogenesis, composition and secretion highlights the heterogeneity of extracellular vesicles. J Cell Sci. 2013 Dec 15;126(Pt 24):5553-65.
7. Rania Ghossoub, et al. Syntenin-ALIX exosome biogenesis and budding into multivesicular bodies are controlled by ARF6 and PLD2. Nat Commun. 2014 Mar 18;5:3477.
8. Caleb M. Richter, et al. Doa4 function in ILV budding is restricted through its interaction with the Vps20 subunit of ESCRT-III. J Cell Sci. 2013 Apr 15; 126(8): 1881–1890.
9. D Michiel Pegtel, et al. Exosomes. Annu Rev Biochem. 2019 Jun 20;88:487-514.
10. Wenchong Zou, et al. Exosome Release Is Regulated by mTORC1. Adv Sci (Weinh). 2018 Dec 11;6(3):1801313.
11. Marina Vietri, et al. The many functions of ESCRTs. Nat Rev Mol Cell Biol. 2020 Jan;21(1):25-42.
12. Matias Ostrowski, et al. Rab27a and Rab27b control different steps of the exosome secretion pathway. Nat Cell Biol. 2010 Jan;12(1):19-30; sup pp 1-13.



