
ҷCуwҝ№ІЎ¶ҫГвТЯөДЧчУГФӯАн
З°СФ
ІЎ¶ҫКЗҢЈРФөДјҡ°ығИјДЙъЈ¬РиТӘФЪЛЮЦчғИҸНЦЖЈ¬ІўҙЩЯMЛьӮғӮчІҘөҪЖдЛыөШ·ҪЎЈФЪИЛоҗЦРЈ¬ҙу¶а”өЕRҙІПакPёРИҫҒнЧФЖдЛы„УОпЈ¬Я@Т»Я^іМИФФЪА^АmЎЈЧоҪьөДАэЧУ°ьАЁИЛоҗГвТЯИұПЭІЎ¶ҫЈЁHIVЈ©Ўў°ЈІ©АӯІЎ¶ҫЎўSARSІЎ¶ҫЈ¬ZIKAІЎ¶ҫЈ¬РВРН№Ъ оІЎ¶ҫөИЎЈ
ІЎ¶ҫёРИҫәЬЙЩКЗЦұҪУЦВГьөДЈ¬јҙК№ЛьӮғҢҰҶОӮҖјҡ°ыҫЯУРёЯ¶ИөДјҡ°ыИЬҪвРФЎЈ
ЛАНцВКНЁіЈ°lЙъФЪІЎ¶ҫОп·NйgМшЬSЎўІЎ¶ҫҪӣҡvЦШҙуҝ№ФӯЧғ»ҜЈЁјҙБчёРІЎ¶ҫЈ©»тЛЮЦчГвТЯКЬ“p•rЎЈ
ІЎ¶ҫЯMИлј°ёРИҫ
ҷCуwҪMҝ—®aЙъҙу¶а”өІЎ¶ҫөДЯMИләНёРИҫөДХПөKЎЈЖдЦРЧоУРР§өДКЗЖӨДwәНХіДӨұнГжМṩөДҷCРөЖБХПЈ¬ТФј°ДcөАөД»ҜҢW”іҢҰӯhҫі ЎЈ
ФS¶аіЈТҠөДИЛоҗІЎ¶ҫІЎФӯуwНЁЯ^ОёДcөАЯMИлЈ¬°ьАЁЭҶ оІЎ¶ҫЎўДcПЩІЎ¶ҫәНјЧРНёОСЧІЎ¶ҫЈЁHAVЈ©ЎЈЯ@Р©НЁіЈКЗНЁЯ^ИЛЕcИЛөДҪУУ|»тКЬОЫИҫөДКіОпәНЛ®ӮчІҘөДЎЈ
УЙБчёРІЎ¶ҫЎўұЗІЎ¶ҫЎў№Ъ оІЎ¶ҫЎўВйХоІЎ¶ҫЎўЛ®¶»-Һ§ о°’ХоІЎ¶ҫЈЁVZVЈ©әНәфОьөАәП°ыІЎ¶ҫЈЁRSVЈ©ТэЖрөДәфОьөАёРИҫНЁіЈНЁЯ^ҡвИЬДzпwДӯӮчІҘЈ¬ТФј°ИЛЕcИЛЦ®йgөДҪУУ|ӮчІҘЎЈ
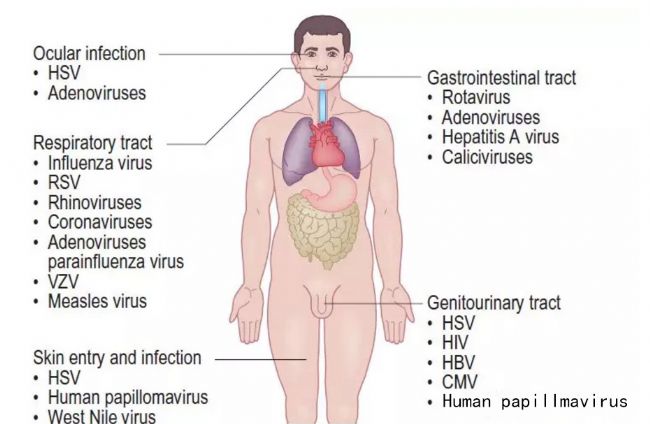
ФS¶а°’ХоІЎ¶ҫбҳҢҰЖӨДw»тХіДӨЈ¬ИзҶОјғ°’ХоІЎ¶ҫЈЁHSVЈ©әНVZVЎЈУИЖдКЗHSVЈ¬ҝЙТФНЁЯ^РЎөДёоӮыәНІБӮыёРИҫҝЪЗ»әНЙъЦіЖчХіДӨЎўСЫҫҰәНЖӨДwЎЈЖдЛы°’ХоІЎ¶ҫЈ¬ИзEpstein-BarrІЎ¶ҫЈЁEBVЈ©әНҫЮјҡ°ыІЎ¶ҫЈЁCMVЈ©Ј¬ТФХіДӨһй°РьcЎЈCMVТІҝЙТФҸДДёУHҙ№ЦұӮчІҘөҪӢлғәЈ¬әЬЙЩНЁЯ^Э”СӘЎЈИЛИйо^БцІЎ¶ҫЈЁHPVЈ©бҳҢҰЖӨДwәНХіДӨЈ¬Ң§ЦВраЈ¬ІўҝЙДЬЮD»Ҝјҡ°ыЈ¬ХT°l°©°YЈ¬ИзҢmоi°©ЎЈ
Т»Р©ІЎ¶ҫЈ¬ИзОчДбБ_ІЎ¶ҫЎўөЗёпҹбІЎ¶ҫЎў°лБЦІЎ¶ҫәНэRҝЁІЎ¶ҫЈ¬ҝЙТФНЁЯ^АҘПxГҪҪйЯMИлЖӨДwЎЈ
°¬ЧМІЎ¶ҫәНТТРНёОСЧІЎ¶ҫЈЁHBVЈ©НЁіЈНЁЯ^РФҪУУ|ӮчІҘЎЈ°¬ЧМІЎ¶ҫЎўHBVәНұыРНёОСЧІЎ¶ҫЈЁHCVЈ©ТІҝЙТФНЁЯ^Э”СӘ»тКЬОЫИҫөДбҳо^ЦұҪУЯMИлСӘТәӮчИҫҪoИЛоҗЎЈ
ҙу¶а”өИЛоҗІЎ¶ҫЦ»ФЪДіР©°РҪMҝ—ЦРҸНЦЖЈ¬ЦчТӘУЙІЎ¶ҫКЬуw·ЦІјөДӣQ¶ЁЎЈФS¶аІЎ¶ҫК№УГғЙ·NКЬуwЈ¬ИзCD4№ІКЬуwәНЪ…»ҜТтЧУКЬуwCCR5ФЪHIVTјҡ°ыЙПөДұнЯ_ЎЈ
ФЪёҪЦшөҪјҡ°ыКЬуwәуЈ¬ІЎ¶ҫҝЙДЬЕcјҡ°ыДӨИЪәП»тғИНМЈ¬И»әуНЁЯ^ЕcДТЕЭДӨЈЁ°ьДӨІЎ¶ҫЈ¬ИзHSVәНHIVЈ©ИЪәПЯMИлјҡ°ыЩ|»тјҡ°ыәЛЈ¬»тФЪјҡ°ыЩ|ЦРТ»ҙОҝзјҡ°ыДӨ»тХTҢ§ғИНМДТЕЭБСҪвЈЁҹoәЛІЎ¶ҫЈ¬ИзNorwalkІЎ¶ҫәНј№Ли»ТЩ|СЧІЎ¶ҫЈ©ЎЈ
И»әуЈ¬ІЎ¶ҫАыУГЛЮЦчјҡ°ыҷCЦЖәНҢЈйTөДІЎ¶ҫҫҺҙaө°°ЧФЪјҡ°ығИҝмЛЩҸНЦЖЎЈТ»ө©ЛьӮғФЪјҡ°ығИ·ұЦіЈ¬ФS¶аІЎ¶ҫҫН•юХTҢ§јҡ°ыИЬҪвЈ¬ТФҙЩЯMРВөДӮчИҫРФІЎ¶ҫЈЁАэИзЈ¬¶»ІЎ¶ҫЎўј№Ли»ТЩ|СЧІЎ¶ҫәН°’ХоІЎ¶ҫЈ©өДбҢ·ЕЎЈ
ФЪјҡ°ыІ»ЛАНцөДЗйӣrПВЈ¬ЖдЛыІЎ¶ҫНЁЯ^јҡ°ыДӨҸДКЬёРИҫөДјҡ°ыЦРбҢ·ЕіцҒнЈЁАэИз°¬ЧМІЎ¶ҫәНБчёРІЎ¶ҫЈ©ЎЈИ»¶шЈ¬ІЎ¶ҫЯMИлИЛуwә󣬕юУцөҪФS¶аПИМм·АУщЈ¬ІўјӨ»оЯm‘ӘРФГвТЯЎЈ
НЁЯ^ТЯГзЈ¬іЙ№ҰАыУГЯ@Р©·АУщКЦ¶ОЈ¬ИФИ»КЗ‘р„ЩФS¶аІЎ¶ҫёРӮчИҫјІІЎөДәЛРД·Ҫ·ЁЈ¬МШ„eКЗДЗР©Ң§ЦВВэРФёРИҫөДІЎ¶ҫЈ¬ИзHIVәНHCV.
МмИ»ГвТЯ
ІЎ¶ҫёРИҫТэЖрЛЮЦчҸV·әөД·АУщҷCЦЖЈ¬ПИМм·АУщҝЙТФЧиЦ№»тТЦЦЖіхЖЪёРИҫЈ¬ұЈЧoјҡ°ыГвКЬёРИҫЈ¬»тПыңзІЎ¶ҫёРИҫөДјҡ°ыЎЈ
ПИМмҷCЦЖФзФЪЯm‘ӘРФГвТЯөДР§‘ӘЧғөГ»оЬSЦ®З°ҫНТСіц¬FЈ¬ө«ЛьӮғҢҰУЪНЁЯ^јӨ°lҙЩЯMГвТЯјҡ°ы»о»ҜөДСЧ°YЈ¬ҒнҶў„УЯm‘ӘРФГвТЯКЗЦБкPЦШТӘөДЎЈ
ПИМмГвТЯ·АУщКЗНЁЯ^ДЈКҪЧR„eКЬуwЈЁPRRSЈ©Ҷў„УөДЈ¬ЛьЧR„eІЎФӯуwПакP·ЦЧУДЈКҪЈЁPAMPЈ©Ј¬°ьАЁTollҳУКЬуwЈЁTLRЈ©јТЧеөДҝзДӨКЬуwЎўғЙӮҖјҡ°ығИКЬуwјТЧеЈЁNODҳУКЬуwЈЁNLRsЈ©әНRIG-IҳУҪвРэГёЈЁRLHsЈ©Ј©Ј¬ТФј°әЪЙ«ЛШБц-2ЈЁAIM2Ј©ЦРИұК§өДӮчёРЖч·ЦЧУЎЈ
ҙЛНвЈ¬ӯhшBЬХТ»БЧЛб-ПЩЬХТ»БЧЛбәПіЙГёcGASЎўDDX41ЎўIFI16әНz-DNAҪYәПө°°Ч1ЈЁZBP1Ј©·ЦЧУҝЙТФёРЦӘјҡ°ыЩ|DNAЎЈ
Я@Р©јҡ°ыӮчёРЖчҙЩЯMIL-1Ј¬IL-18ЎўIРНЈЁα/β)-IFNЈ©ЎўёЙ”_»щТтәНСЧ°Yјҡ°ыТтЧУј°Ъ…»ҜТтЧУөДұнЯ_ЎЈTLRsКЗУЙФS¶ајҡ°ыұнЯ_өДјҡ°ыұнГж»тғИРЎуwДӨҪYәПө°°ЧЈ¬°ьАЁҳдН» ојҡ°ыЈЁDCSЈ©ЎўҫЮКЙјҡ°ыЎўБЬ°Нјҡ°ыәНҢҚЩ|јҡ°ыЎЈ
ФЪҙу¶а”өјҡ°ыоҗРНЦРЈ¬TLRsөДұнЯ_ФЪәЬҙуіМ¶ИЙПКЗҝЙХTҢ§өДЈ¬Т»Р©ЈЁTLR7/8/9Ј©ФЪҢЈйTөДқ{јҡ°ыҳУDCSёЯЛ®ЖҪЙПұнЯ_Ј¬ТФҝмЛЩ®aЙъIFNЎЈ
І»Н¬өДTLR·ЦЧУЧR„eМШ¶ЁөДІЎ¶ҫ®aОпЈ¬ИзҶОжңәНлpжңRNAЈЁTLR3әНTLR7/8Ј©»тлpжңDNAЈЁTLR9Ј©Ј¬ИзПВҲDЈә

NLRPsКЗPAMPSөДөЪ¶юоҗ°ық{ӮчёРЖчЈ¬НЁЯ^ЯmЕдЖчө°°ЧASCјӨ»оСЧ°YРЎуwЎЈЯ@Р©°ьАЁNLR PЈЁ»тNALPЈ©ЎўNODәНIPAF/NAIPКЬуwЎЈУРИэ·NЦчТӘөДСЧ°Yуw…ўЕcБЛҝ№ІЎ¶ҫГвТЯЈәNLRP3СЧ°YуwЎўRIG-IСЧ°YуwәНAIM2СЧ°YуwЎЈПИМм·АУщПөҪyУЙ¶а·Nјҡ°ыіЙ·ЦәНФS¶аМШКвө°°ЧЩ|ҪMіЙЎЈТСЦӘәНСРҫҝөГЧо¶аөДҝ№ІЎ¶ҫө°°ЧКЗα/βIFNsЈ¬ЛыӮғНЁЯ^IРНIFNКЬуw°l“]ЧчУГЈ¬ІўТэЖр100¶а·N»щТтЧғ»ҜЎЈЯ@·N“ҝ№ІЎ¶ҫ о‘B”өДәу№ыЦ®Т»КЗТЦЦЖјҡ°ыө°°ЧЩ|әПіЙәН·АЦ№ІЎ¶ҫҸНЦЖЎЈ
¶аӮҖ°Чјҡ°ыҒҶИә…ўЕcПИМм·АУщЈ¬°ьАЁҫЮКЙјҡ°ыЎўDCSЎўЦРРФБЈјҡ°ыЎўЧФИ»ҡўӮыјҡ°ыЈЁNKЈ©ЎўЧФИ»ҡўӮыTјҡ°ыЈЁNKTјҡ°ыЈ©әНγδTјҡ°ыЎЈҙЛНвЈ¬ҪMҝ—јҡ°ыЈ¬°ьАЁіЙАwҫSјҡ°ыЎўЙПЖӨјҡ°ыәНғИЖӨјҡ°ыЈ¬НЁЯ^®aЙъПИМмјҡ°ыТтЧУЈ¬°ьАЁIFN-IәНIL-1Ј¬ұнЯ_PRRSІўҢҰІЎ¶ҫёРИҫЧчіц·ҙ‘ӘЎЈINF-IКЗПИМмГвТЯПөҪyәНЯm‘ӘРФГвТЯПөҪyЦ®йgөДкPжIВ“ПөНЁЯ^јӨ»оDCSәНTјҡ°ыЈ¬ТФј°ұЈЧoTјҡ°ыГвКЬNKјҡ°ыҪйҢ§өД№Ҙ“фЎЈёЙ”_ЛШ-IЯҖДЬјӨ»оNKјҡ°ыІўХTҢ§ЖдЛыҙЩЯMNK·ҙ‘ӘөДјҡ°ыТтЧУЈ¬ИзIFN-γәНIL-12ЎЈNKјҡ°ы®aЙъҙЩСЧјҡ°ыТтЧУЈ¬ҝЙТФҡўЛАёРИҫјҡ°ыІўЕcDCSПа»ҘЧчУГЈ¬КЗөЦУщІЎ¶ҫөДЦШТӘҪMіЙІҝ·ЦЎЈ
NKјҡ°ыҢҰРЎКуәНИЛоҗCMVөДҝ№РФәЬЦШТӘЈ¬ҝЙДЬҢҰHIVЎўБчёРІЎ¶ҫәН°ЈІ©АӯІЎ¶ҫТІәЬЦШТӘ ЎЈНЁЯ^ҡўЛАCD4Tјҡ°ыәНйgҪУХ{№қјҡ°ы¶ҫРФTБЬ°Нјҡ°ыЈЁCT LЈ©·ҙ‘ӘЈ¬NKјҡ°ыТІҝЙТФҝШЦЖҢҰІЎ¶ҫЯm‘ӘРФГвТЯ·ҙ‘ӘЎЈNKTјҡ°ыҝЙДЬҢҰДіР©ІЎ¶ҫЈ¬ИзБчёРІЎ¶ҫЈ¬МṩһЩҝ№ФӯМШ®җРФөДМмИ»ГвТЯұЈЧoЎЈ
ҺЧоҗМмИ»ЛЮЦчө°°ЧФЪҝ№ІЎ¶ҫ·АУщЦРЖрЧчУГЎЈЖдЦР°ьАЁМмИ»ҝ№уwЈ¬Я@·Nҝ№уwҝЙДЬФЪ·АУщДіР©ІЎ¶ҫёРИҫ·ҪГжЖрЧчУГЈ¬ЯҖ°ьАЁpentraxinsәНСaуwө°°ЧЎЈУРР©ІЎ¶ҫҝЙДЬұ»јӨ»оСaуwЦұҪУңз»оЈ¬»тұ»ҪYәПәННМКЙБЛСaуwҪYәПІЎ¶ҫөДНМКЙјҡ°ыЖЖүДЎЈІЎ¶ҫёРИҫХTҢ§өДҺЧ·NҙЩСЧјҡ°ыТтЧУәНЪ…»ҜТтЧУФЪ·АУщЦРТІЖрЦшкPжIЧчУГЎЈЖдЦРЧоЦчТӘөДКЗIL-1әНIL-1јТЧеөДЖдЛыіЙҶTЈ¬°ьАЁIL-18әНIL-33ЎЈ
Я@Р©јҡ°ыТтЧУУ°н‘ПИМмәНЯm‘ӘРФГвТЯјҡ°ыЈ¬ІўФЪҝ№ІЎ¶ҫ·АУщЦР°l“]кPжIЧчУГЎЈЖдЛыҝ№ІЎ¶ҫјҡ°ыТтЧУФЪёРИҫәуФзЖЪ®aЙъЈ¬ИзTNF-αЎўIFN-γЎўIL-12ЎўIL-6әНЪ…»ҜТтЧУЈ¬ИзMIP-1αЎЈ
МШ„eКЗЈ¬IL-12КЗNKјҡ°ыЦРIFN-γөДУРР§ХTҢ§„©.СЧ°YЪ…»ҜТтЧУТІҝЙДЬНЁЯ^ФЪёРИҫІҝО»…fХ{ҫЮКЙјҡ°ыЎўЦРРФБЈјҡ°ыЎўDCәНNKјҡ°ыөД·ҙ‘ӘЈ¬ФЪПИМмҝ№ІЎ¶ҫ·АУщЦР°l“]ЦШТӘЧчУГЎЈ
І»ғHПИМмГвТЯөДЯ@Р©іЙ·Ц…ўЕcБЛҢҰІЎ¶ҫөДіхІҪұЈЧoЈ»ҺЧӮҖіЙ·ЦЈЁАэИзPPRsЈ»јҡ°ыТтЧУIFN-IЎўIL-1ЎўIL-33әНIL-12Ј»ТФј°НМКЙјҡ°ыЈ¬°ьАЁҫЮКЙјҡ°ыЎўҶОәЛјҡ°ыәНDCsЈ©УРЦъУЪЛЬФмлSәуҢҰІЎ¶ҫІЎФӯуwөДЯm‘ӘРФ·ҙ‘ӘөДРФЩ|әНУРР§РФЎЈ
АэИзЈ¬DCSРиТӘПИМмРЕМ–Ј¬ИзIFN-IәНIL-12Ј¬ІЕДЬіЙКмәН®aЙъЧојСөДTјҡ°ыјӨ»о.ҙЛНвЈ¬н‘‘ӘІЎ¶ҫөДCD8Tјҡ°ыРиТӘIFN-IәНIL-33РЕМ–Ғн”UХ№әНРОіЙУӣ‘ӣЎЈ
ІЎ¶ҫёРИҫТэЖрөДПИМм·ҙ‘ӘөДҙуРЎәНоҗРН¶јҢҰЯm‘ӘРФГвТЯ·ҙ‘ӘөД®aЙъУРЦшГчп@өДУ°н‘ЎЈ

ПИМмГвТЯНЁіЈЦ»•юңpҫҸ¶шІ»КЗНЈЦ№ІЎ¶ҫёРИҫЈ¬ҪoЯm‘ӘРФГвТЯ·ҙ‘Ә„“Фм•rйgЎЈ
Яm‘ӘРФГвТЯөДғЙӮҖЦчТӘІҝ·ЦЈ¬ҝ№уwҪйҢ§әНTјҡ°ыҪйҢ§Ј¬ЦчТӘбҳҢҰІ»Н¬өД°Рьc.
ҝ№уwНЁіЈНЁЯ^ЕcУОлxөДІЎ¶ҫоwБЈҪYәП¶ш°l“]ЧчУГЈ¬Чи”аЛЮЦчјҡ°ыөДёРИҫ ЎЈ
Па·ҙЈ¬Tјҡ°ыөДЧчУГЦчТӘКЗЧR„eәНЖЖүДІЎ¶ҫёРИҫөДјҡ°ыЈ¬»тҶў„УСЧ°Y·ҙ‘ӘЎЈУЙУЪЛщУРІЎ¶ҫ¶јФЪјҡ°ығИҸНЦЖЈ¬¶шЗТФS¶аІЎ¶ҫҝЙТФФЪјҡ°ыйgЦұҪУӮчІҘЈ¬¶шІ»•юЦШРВЯMИлјҡ°ыНвӯhҫіЈ¬ТтҙЛёРИҫөДҪвӣQёь¶аөШТАЩҮУЪTјҡ°ы№ҰДЬ¶шІ»КЗҝ№уw№ҰДЬ ЎЈИ»¶шЈ¬ҸV·әЦРәНҝ№ІЎ¶ҫҝ№уwҫЯУРУРР§ЦОҜҹФS¶аІ»Н¬ИЛоҗёРИҫөДқ“БҰЈ¬°ьАЁHIVЎўБчёРІЎ¶ҫәН°ЈІ©АӯІЎ¶ҫЎЈ
ЧоҪьөДЯMХ№К№СРҫҝИЛҶTДЬүт·ЦлxәНиb¶ЁИЛоҗбҳҢҰЯ@Р©ІЎФӯуwәНЖдЛыІЎФӯуwөДҶОҝЛВЎҝ№уwЈ¬һйРВҜҹ·ЁМṩБЛЗ°ҫ°Ј¬ІўһйТЯГзФOУӢМṩБЛЦШТӘөДТҠҪв.ҝ№ІЎ¶ҫҝ№уwЧчһйҝ№ФЩёРИҫөДГвТЯұЈЧoЖБХПТІКЗ·ЗіЈЦШТӘөДЎЈЦчТӘКЗИлҝЪМҺЈЁНЁіЈКЗХіДӨұнГжЈ©УРҝ№уwЈ¬Я@ЕcБчёРЎўHSVәН°¬ЧМІЎ¶ҫёРИҫМШ„eПакP.И»¶шЈ¬ИзәО®aЙъДЬТэЖрЧојСҝ№уw·ҙ‘ӘөДТЯГзЈ¬°ьАЁҸV·әөДЦРәНҝ№уwЈ¬ИФИ»КЗТ»ӮҖЙРОҙҪвӣQөДЦШТӘҶ–о}ЎЈ
Яm‘ӘРФГвТЯөДҶў„УГЬЗРТАЩҮУЪјӨ»оҝ№ФӯМбіКјҡ°ыЈЁAPCЈ©өДФзЖЪПИМмҷCЦЖЈ¬ЦчТӘКЗDCsЎЈНЁЯ^Ъ…»ҜТтЧУәНјҡ°ыТтЧУРЕМ–ҢўAPCәНБЬ°Нјҡ°ыОьТэөҪБЬ°НҪMҝ—ЦРЈ¬ІўФЪДЗАпұЈБфҺЧМмЈ¬ТФҙЩЯMУРР§өДјҡ°ыйgПа»ҘЧчУГЎЈ
ҙОјүБЬ°НҪMҝ—өДҪYҳӢНЁЯ^Ц§іЦРФ»щЩ|јҡ°ыҫWҪjәНҫЦІҝЪ…»ҜТтЧУМЭ¶ИЦ§іЦЯm‘ӘРФГвТЯПөҪyјҡ°ыЦ®йgөД…fХ{Па»ҘЧчУГЎЈ
ХTҢ§КВјю°lЙъФЪБЬ°НҪYТэБчёРИҫІҝО»»тЖўЕKЈ¬Из№ыІЎ¶ҫЯMИлСӘТәЎЈІЎ¶ҫҝ№ФӯӮчЯfөҪБЬ°НҪYНЁіЈ°lЙъФЪDCSЦРЎЈУРР©ІЎ¶ҫ•ю“pәҰAPCөД№ҰДЬЈ¬ИзHSVәНВйХоІЎ¶ҫЈ¬ҸД¶шТЦЦЖDCөДіЙКмЎЈ
ФЪЖўЕKәНБЬ°НҪYЈ¬Bјҡ°ыһVЕЭЈ¬Tјҡ°ы…^Ј¬Bјҡ°ыУцөҪҝ№ФӯәујӨ»оЎЈТ»Р©»о»ҜөДBјҡ°ыіЙһй¶МүЫГьөДқ{јҡ°ыЈ¬¶шБнТ»Р©„tТЖ„УөҪBјҡ°ыВСЕЭөДЯ…ҫүЈ¬ІўНЁЯ^ФЪBјҡ°ыMHCIIоҗ·ЦЧУЙПіКЯfҝ№ФӯлДЕcҝ№ФӯМШ®җРФЭoЦъCD4Tјҡ°ыПа»ҘЧчУГЎЈ
Я@Р©BCL6ТАЩҮөДCD4TһVЕЭЦъ„©ЈЁTfhЈ©јҡ°ыКЗҢЈйTһйBјҡ°ы·ҙ‘ӘМṩҺНЦъөДјҡ°ыЈ¬КЗҙЩЯMәНХ{№қBјҡ°ы·ҙ‘ӘЛщұШРиөДЎЈ
»о»ҜөДBјҡ°ыФЪCD4Tfhјҡ°ыөДҺНЦъПВҶў„УЙъ°lЦРРДЈЁGCЈ©·ҙ‘ӘЈ¬ҙ_ұЈуwјҡ°ыөДёЯН»ЧғәНУHәНБҰіЙКмЈ¬ТФЯx“сёЯУHәНБҰЎў®aЙъҝ№уwЎўйLүЫГьөДқ{јҡ°ыТФј°Уӣ‘ӣBјҡ°ыЎЈ
ФЪ·ЦЧУЛ®ЖҪЙПЈ¬ЮDдӣТтЧУBlimp-1ЎўXBP-1әНIRF-4өДЙПХ{ӣQ¶ЁБЛқ{јҡ°ыөДРОіЙЈ¬¶шPax-5өДұнЯ_„tГиКцБЛЧў¶ЁТӘЯMРРGC·ҙ‘ӘөДBјҡ°ыәНУӣ‘ӣBјҡ°ыЧVПөЎЈ
ҝ№уwҪYәПұнЯ_ФЪМмИ»ө°°ЧЩ|ұнГжөДұнО»Ј¬ЧиЦ№ІЎ¶ҫёҪЦш»тқBНёөҪДҝҳЛјҡ°ыЎЈЦ®әуНЁЯ^CDC»тХЯADCCөДЧчУГЈ¬БСҪвІЎ¶ҫЎЈЕј –Ј¬ҝ№уwҪYәПІЎ¶ҫөДFCКЬуwҪYәПҝЙДЬҢ§ЦВёРИҫЈ¬ІўҢ§ЦВёьҮАЦШөДҪMҝ—“pӮыЈ¬°lЙъФЪөЗёпҹбәНHIVёРИҫЎЈ…ўЕcұЈЧoИЛуwХіДӨұнГжөДҝ№уwЦчТӘКЗ·ЦГЪРФГвТЯЗтө°°ЧAЈЁIgAЈ©Ј¬ө«СӘЗеСЬЙъөДIgGТІҝЙДЬҫЯУРұЈЧoЧчУГЈ¬МШ„eКЗФЪкҺөАХіДӨөИІҝО» ЎЈ
Я@ғЙ·Nҝ№уwН¬РНЦчТӘЧчУГУЪЧи”аЙПЖӨјҡ°ыөДёРИҫЈ¬ұM№ЬФЪДіР©ЗйӣrПВЈ¬ҝ№уwҝЙТФҢўҝ№ФӯҸДуwғИНЁЯ^ЙПЖӨјҡ°ыЮDЯ\өҪуwНв ЎЈ
ЕcСӘЗеҝ№уwПаұИЈ¬ХіДӨҝ№уwөДіЦАm•rйgТӘ¶МөГ¶аЈ¬Я@ФЪТ»¶ЁіМ¶ИЙПҪвбҢБЛһйКІГҙҢҰХіДӨІЎФӯуwөДГвТЯНЁіЈұИҢҰИ«ЙнІЎ¶ҫёРИҫөДГвТЯ•rйg¶МөГ¶аЎЈ
ҝӮҪYҝ№ёРИҫГвТЯИзПВұн
ГвТЯУӣ‘ӣКЗЯm‘ӘРФГвТЯөДЦчТӘМШХчЎЈТЯГзҢWөДДҝөДКЗХTҢ§йLЖЪГвТЯУӣ‘ӣЈ¬ТФ·АЦ№ФЩёРИҫ ЎЈФЪёРИҫДіР©ІЎ¶ҫәуЈ¬Уӣ‘ӣҝЙДЬ•ю®җіЈйLүЫЈ¬ҝЙДЬҢҰЛЮЦчөДЙъГьУРУ°н‘(ьSҹбІЎәНМм»ЁІЎ¶ҫ Ј©ЎЈ
Уӣ‘ӣөД¶ЁБxКЗМШ®җРФБЬ°Нјҡ°ыәН®aЙъҝ№уwөДқ{јҡ°ыөДіЦАmҙжФЪЈ¬¶шІ»КЗҝ№ФӯөДіЦАmҙжФЪЈ¬ТФХTҢ§БЬ°Нјҡ°ыөДіЦАmјӨ»оЎЈ
ҢҰІЎ¶ҫөДуwТәУӣ‘ӣЙжј°№ЗЛиЦРөДйLүЫГьқ{јҡ°ыЈ¬Я@МṩБЛТ»ӮҖіЦАmөДөНЛ®ЖҪСӘЗеҝ№уwҒнФҙЎЈуwТәГвТЯөДҫSіЦТІЙжј°Т»Иә·Җ‘BҫSіЦУӣ‘ӣBјҡ°ыЈ¬Я@ҝЙДЬКЗҫSіЦ·Җ¶Ё”өБҝөДйLүЫГьқ{јҡ°ыЛщұШРиөДЎЈУӣ‘ӣTјҡ°ыіШУЙјҡ°ыТтЧУIL-7әНIL-15ҝШЦЖөДөНЛ®ЖҪ·Җ‘B·ЦБСХ{№қЎЈҢҰУЪУӣ‘ӣуwCD8Tјҡ°ыҒнХfЈ¬IL-7ҢҰУЪҙж»оКЗЦБкPЦШТӘөДЈ¬¶шIL-15ҢҰУЪҫSіЦУӣ‘ӣуwTјҡ°ыіШҙуРЎөДөНЛ®ЖҪФцЦіКЗЦБкPЦШТӘөДЎЈ
Р§‘ӘәНУӣ‘ӣTјҡ°ы·Ц»ҜЎЈҝ№ФӯҙМјӨ”UҙуР§‘Әјҡ°ыЈ¬ЖдЦРҙу¶а”өФЪКХҝsЖЪЛАНцЎЈлSЦш•rйgөДНЖТЖЈ¬РОіЙөДР§‘ӘУӣ‘ӣTЈЁTEMЈ©јҡ°ыЦрқuЮD»ҜһйЦРСлУӣ‘ӣTЈЁTCMЈ©јҡ°ыЈ¬ұнГжҳЛУӣұнЯ_°lЙъПа‘ӘөДЧғ»ҜЎЈТ»Р©Р§‘ӘTјҡ°ы°lХ№һйсvБфУӣ‘ӣTЈЁTRMЈ©јҡ°ыЈ¬Я@Р©јҡ°ыіЦАmҙжФЪУЪҪMҝ—ЦРЈ¬І»ФЩЯMИлСӯӯhЎЈ
ІЎ¶ҫГвТЯМУұЬөДҷCЦЖ
ФS¶аІЎ¶ҫЈ¬ІЙУГңpИхГвТЯ»тСУЯtөДІЯВФҒнТҺұЬГвТЯПөҪyөДёч·N·АУщҷCЦЖЈ¬К№ЛьӮғУР•rйgЯMТ»ІҪҸНЦЖ»тМУұЬұOңyЎЈ
ҫЯуwҷCЦЖј°ҢҚАэТҠПВұн
ФuЈә
SARSЈ¬H7N9Ј¬H5N1, 2019-nCov, MERs......ІЎ¶ҫЛБЕ°Ј¬І»”аҪoИЛоҗМбРСЈ¬Ң‘ьc»щөAөД–|ОчЈ¬№©ҙујТЯ@ғЙМмФЪјТҹoБД•rҝҙҝҙЎЈ
ТЯГзй_°lЈ¬ҝЙТФ…ўҝјЦ®З°ОДХВЈә·ҙПтТЯГзҢW2.0Йэјү°жЈә»щУЪҝ№ФӯҪYҳӢөДТЯГзФOУӢј°°ёАэЎЈ
РВРН№Ъ оІЎ¶ҫ·АЦОХҲйҶЧxҮшјТ№Щ·ҪЦёДПЎЈ
…ўҝјОД«I
-
Kanneganti TD. Central roles of NLRs and inflammasomes in viral infection. Nat Rev Immunol 2010;10:688–98.
-
Wherry EJ, Kurachi M. Molecular and cellular insights into T cell exhaustion. Nat Rev Immunol 2015;15:486–99.
-
Mueller SN, Gebhardt T, Carbone FR, et al. Memory T cell subsets, migration patterns, and tissue residence. Annu Rev Immunol 2013;31:137–61.
-
Barry T. Rouse, Scott N. MuellerЈ¬Host Defenses to VirusesЈ¬Clinical immunologyЈ¬Book • 5th Edition • 2019

- H5N1ЗЭБчёРІЎ¶ҫёРИҫпLлUЕcФзЖЪҷzңy·Ҫ·Ё
- CDR3№ӨіМЕcЦРәНҝ№уw¶ЁЦЖЈәұЈЧCРНҝ№уwЦШЛЬЦОҜҹРВҳЛңК
- ИзәОҳӢҪЁёЯР§Н»ЧғГёОДҺмЈҝ
- ҝ№уw¶ЁЦЖЎўј{ГЧҝ№уwЕcЕдҢҰҝ№уwөД„“РВБҰБҝ
- КЙҫъуwХ№КҫјјРgЈәй_ҶўЙъОпбtЛҺРВјoФӘөДиҖіЧ
- ИэҙујјРg…fН¬Ј¬Ҫвжiҝ№уwСРҫҝРВОҙҒн
- ¶алДОДҺмҳӢҪЁЕcәYЯx·ю„ХЈәјУЛЩЛҺОпСР°lөДәЛРДтҢ„УБҰ
- КЙҫъуwҪЁҺмЕcҝ№уwКЙҫъуwОДҺмөД…fН¬„“РВ
- СРУ‘•юЈәAIтҢ„УЩ|ЧVјјРgЩxДЬЙъОпЛҺСР°lЕcГвТЯЦОҜҹ
- °ЩҠWЩҗҲDСыДъПајsChinaBioәПЧчХ“үҜ
- °ЩҠWЩҗҲDЕcMyricxЯ_іЙҝ№уwФu№А…fЧhУЪADCЛҺОпй_°l
- 2023Дк°ЩЭиЙъОпҝЖјјХ\Жё¶аӮҖҚҸО»
- ЧЈЩR°ЩЭиЙъОпЙПәЈСР°lЦРРДҶМЯwРВЦ·ЕcРВ№ЩҫWЙПҫҖ
- қъ·ІЙъОпББПаINDёРИҫРФјІІЎПИЯM·ЦЧУФ\”а‘ӘУГХ“үҜ
- өvәНбtЛҺҝ№уwФЪИ«ЗтIЖЪЕRҙІСРҫҝНкіЙКЧАэ»јХЯҪoЛҺ
- °ЩҠWЩҗҲDЕcДПҫ©ХэҙуМмЗзәһКрИ«ИЛҝ№уwәПЧчй_°l…fЧh


